
M.T. Sakr
Department of Botany Agriculture, Faculty Agriculture, Mansoura University, 35516, Egypt
M.A. El-Metwally
Agriculture Research Center, Department of Mycol Research and Plant Dis. Surv., Plant Pathology Research Institute, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 8 | Page No.: 624-630







ABSTRACT
Two field experiments were carried out during the two growing seasons (2005/2006; 2006/2007) to investigate the role of some plant antioxidant materials such as ascorbic acid, glutathione, α-tocopherol and spermine in alleviating the harmful effects caused by soil salt levels (3840 and 6080 mg L-1) on wheat plant. The grains were pre-soaked then the plants sprayed with any of antioxidants used. Moreover, the data showed that 6080 mg L-1 soil salt level alone or in combination with any of applied antioxidants increased the activity of total peroxidase, ascorbic peroxidase, superoxide dismutase and catalase in wheat leaves. In addition, salinity level (6080 mg L-1) alone or in combination with any of applied antioxidants increased the endogenous contents of ascorbic acid and glutathione and total phenols but decreased carotenoids. It could be concluded that salt soil stress depressed all of growth parameters and yield components. The data also revealed that the different antioxidants could partially alleviate the harmful effect of salinity stress which reflected on growth and yield of wheat plant.
PDF Abstract XML References Citation
How to cite this article
M.T. Sakr and M.A. El-Metwally, 2009. Alleviation of the Harmful Effects of Soil Salt Stress on Growth, Yield and Endogenous Antioxidant Content of Wheat Plant by Application of Antioxidants. Pakistan Journal of Biological Sciences, 12: 624-630.
DOI: 10.3923/pjbs.2009.624.630
URL: https://scialert.net/abstract/?doi=pjbs.2009.624.630
DOI: 10.3923/pjbs.2009.624.630
URL: https://scialert.net/abstract/?doi=pjbs.2009.624.630
INTRODUCTION
Soil salinity is one of the major abiotic stress affecting crop growth and productivity. Salt stress causes inhibition of growth and development, reduction in photosynthesis, respiration and protein synthesis and disturbs nucleic acid metabolism (Sairam and Srivastava, 2002). Sakr et al. (2007) indicated that salinity suppressed both cell division and cell enlargement proportionally in wheat plants.
Regarding the effect of antioxidant on wheat under salinity stress. Shalata and Neumann (2001) found that ascorbic acid acts directly to neutralize superoxide radicals, singlet oxygen or superoxide and as a secondary antioxidant during reductive recycling of the oxidized form tocopherol.
It is now widely accepted that Reactive Oxygen Species (ROS) are responsible for various stress-induced damage to macromolecules and ultimately to cellular structure. Consequently the role of antioxidant enzymes viz. superoxide dismutase (SOD), ascorbate peroxidase (APX), Glutathione Reductase (GR) and catalase (CAT), as well as metabolites like ascorbic acid, glutathione, α-tocopherol, flavonoids and carotenoids (CAR) are responsible for the quenching of ROS becomes very important (Sairam and Srivastava, 2002).
Regarding enzymatic antioxidants activity: Vaidyanathan et al. (2003) reported that the non-enzymatic antioxidants (glutathione and ascorbate) showed an accumulation in root tissues in plants subjected to salt stress. Plants must possess efficient antioxidant system such as SOD, APX, GR, DIIAR and CAT as well as metabolites viz., ascorbic acid, glutathione, α-tocopherol, carotenoid and flavanoids etc. (Smirnoff, 1995).
As for the effect of polyamines: (Zhao and Qin, 2004) has established the specific role of polyamines in maintaining a cation-anion balance in plant tissues and by stabilizing membranes at high external salinity. Polyamines had ameliorating effect on all morphological and physiological characters and prevented degradation of chlorophyll, however polyamines enhanced accumulation of all organic compounds under salinity stress study, except phenols. Therefore, the present investigation aimed to improve the productivity of wheat plant grown under soil salt stress by using some external antioxidant.
MATERIALS AND METHODS
Two field experiments were carried out at Seed Technology Unit, Tag El-Ezz Research Station, Dakahlia Governorate, Agric. Res. Center, Ministry of Agric., Egypt during 2005/2006 and 2006/2007 growing seasons to investigate the role of selected antioxidants on mitigation or alleviation the harmful effect of soil salt stress on wheat plant. Wheat (Triticum aestivum) grains (var. Gemeza 9) kindly were supplied by Plant Breeding Section, Agric. Res. Center, Ministry of Agric., Giza, Egypt. Two different soil areas differ in their soil salt stress were chosen. Soil salt stress in the first area equal 850 mg L-1 (1.32 dS m-1 control). While the second equal 3840 mg L-1 (6 dS m-1) and the third salt soil area was 6080 mg L-1 (9.5 dS m-1). Each area was divided into 5 groups represented by the different applied antioxidants. Uniform grains were presoaked for 6 h before sowing in any of antioxidants i.e., ascorbic acid (100 mg L-1), glutathione (100 mg L-1), α-Tocopherol (50 mg L-1), spermine (10 mg L-1) as well as tap water. Uniform presoaked wheat grains were sown on November 15th 2005 in the two different soil salt areas. The wheat plants were sprayed with the same antioxidant concentrations at 30, 60 and 90 days from sowing. Automatic atomizers were used for spraying the applied antioxidants after adding tween-20 as a wetting agent (0.05%). All the normal culture practices of growing wheat plants were applied as usual manner followed by the farmer in the distinct. Growth parameters were measured at vegetative stage (75 days after sowing) at harvesting stage yield and its components were recorded. The leaves were collected after 75 days from sowing for each treatment then frozen and stored till analysis. Each treatment replicated 3 times and arranged in a complete randomized block design. The data of experiments were statistically analyzed as technique of the Analysis of Variance (ANOVA) according to Gomez and Gomez (1984). The treatment means were compared using the Least Significant Differences (LSD).
Biochemical constituents
Non-enzymatic antioxidant contents
Ascorbic acid determination: The 0.5 g of fresh leaves was ground in 50 mL of 2% (w/v) metaphosphoric acid using mortar and pastel and centrifuged for 30 min at 13000 rpm at 4°C. The ascorbate content (μmol g-1 FW) was measured in the supernatant at 25°C. The absorbance of red color was measured at 520 nm according to Omaye et al. (1979).
Total glutathione determination: The level of total glutathione (GSH) was determined in the fresh leaves with Ellman’s reagent according to De Vos et al. (1992). Three hundred microliter of sample buffer were mixed with 630 μL of 0.5 M K2HPO4 and 25 μL of mM 5, 5-dithiobis (2-nitrobenzoic acid) (final pH 7). The absorbance at 412 nm was read after 2 min GSH was used as a standard.
Total phenols determination: One gram of the fresh leaves were macerated in 5-10 mL 80% ethanol for at least 24 h at 0°C, the alcohol was clarified, the remained residue was re-extracted with 5-10 mL 80% ethanol 3 times. At the end, the clarified extract was completed to 50 mL using 80% ethanol. The colorimetric method of Folin-Denis as described by Daniel and George (1972) was employed for the chemical determination of phenolic compounds by reading the developed blue color at 725 nm. using 0.5 mL 80% ethanol and reagents only as a blank.
Carotenoids determination: Leaf samples (0.5 g from the third upper foliage leaf) were subjected to extraction by methanol for 24 h at laboratory temperature after adding a trace from sodium carbonate (Robinson et al., 1983) then carotenoids were determined spectrophotometrically (Spekol11) at wave length 452 and calculated by equation introduced by Mackiny (1941).
Enzymatic antioxidant activity determination:
Ascorbate peroxidase activity determination: Ascorbate Peroxidase (As-POD) was assayed spectrophotochemically according to Fielding (1978). The assay was carried out at 25°C in 1.0 cm light path cuvette and the reaction mixture consisted of 1500 μL phosphate buffer, 20 μL EDTA, 1000 μL sodium ascorbate and enzyme extract (20 μL). After mixing the reaction was initiated by adding the 480 μL H2O2 and decreasing in optical density at 290 nm against blank (without extract) was continuously recorded every minute (2 min).
Total peroxidases activity determination: Peroxidase was assayed spectrophotochemically according to Amako et al. (1994). The assay was carried out at 25°C in l.0 cm light path cuvette and the reaction mixture consisted of 1500 μL phosphate buffer, 1000 μL pyrogallol and 480 μL H2O2 solution. After mixing the reaction was initiated by adding the enzyme extract (20 μL) and the increase in optical density at 430 nm against blank (without extract) was continuously recorded every minute (for 3 min).
Super Oxide Dismutase (SOD) activity determination: Leaf samples were collected in an ice bucket and brought to the laboratory. Leaves were then washed with distilled water and surface moisture was wiped out. Leaf samples (0.5 g) were homogenized in ice cold 0.1 M phosphate buffer (pH 7.5) containing 0.5 mM EDTA with pre-chilled pestle and mortar. The homogenate was transferred to centrifuge tubes and was centrifuged at 4°C in Beckman refrigerated centrifuge for 15 min at 15000 x g. The supernatant was transferred to 30 mL tubes and referred to enzyme extract.
SOD activity was estimated by recording the decrease in absorbance of superoxide-nitro blue tetrazolium complex by the enzyme. About 3 mL of reaction mixture containing 0.1 mL of 1.5 M sodium carbonate, 0.2 mL of 200 mM methionine, 0.1 mL of 2.25 mM Nitro-blue tetrazolium, 0.1 mL of 3 mM EDTA, 1.5 mL of 100 mM potassium phosphate buffer, 1 mL distilled water and 0.05 mL of enzyme were taken in test tubes in duplicate from each enzyme sample. Two tubes without enzyme extract were taken as control. The reaction was started by adding 0.1 mL riboflavin (60 mM) and placing the tubes below a light source of two 15 W florescent lamps for 15 min reaction was stopped by switching off the light and covering the tube with black cloth. Tubes without enzyme developed maximal color. A non-irradiated complete reaction mixture which did not develop color served as blank. Absorbance was recorded at 560 nm and one unit of enzyme activity was taken as the quantity of enzyme which reduced the absorbance reading of samples 50% of comparison with types lacking enzymes (Dhindsa et al., 1981).
Catalase activity was determined by measuring the rate of H2O2 conversion to O2 at room temperature using a lipid-phase O2 electrode (Hansatech, Norfolk, UK). Approximately 0.5 g of plant tissue, consisting of the apical region of the shoot including the cotyledons, was extracted in 1.5 mL of 0.1 mM Hepes/KOH buffer (pH 7.4)
and then centrifuged at 10000 g for 5 min. The rate of O2 production was measured by adding 50 μL of the supernatant to 0.1 M Hepes (pH 7.4) containing 530 mM H2O2. Catalase activity was calculated on a fresh weight basis to keep the data uniform with the H2O2 measurements and to reduce the chances of distribution as a result of protein synthesis alteration due to heat shock (Vierling, 1991; Bettany, 1995).
RESULTS
Plant growth: The data in Table 1 show that any of applied antioxidants (Ascorbic acid, Glutathione, α-Tocopherol, Spermine) significantly increased dry matter accumulation in stem and leaves of wheat plant compared with untreated plants in the two soil salt areas (A1 and A2) throughout the two growing seasons (2005/2006; 2006/2007). The data also show that applied antioxidants were more effective in soil salt area (A1).
It could be concluded that, the different applied antioxidant materials could counteract the harmful effect of high soil salt stress levels on growth of wheat plant. However, ASA was more effective in this respect.
Yield and its components: The data in Table 1 shows that applied antioxidants significantly increased yield and its components (as spike length, spike weight, number of spikes/plant, number of grains/spike, grains weight/spike and grains yield/plant) in the two salt soil areas especially (A1) compared with the untreated plants throughout the two growing seasons.
| Table 1: | Effect of exogenous applied antioxidants on growth and yield of wheat plant grown under high soil salt stress conditions (3840 and 6080 mg L-1), during the two growing seasons (2005/2006; 2006/2007) |
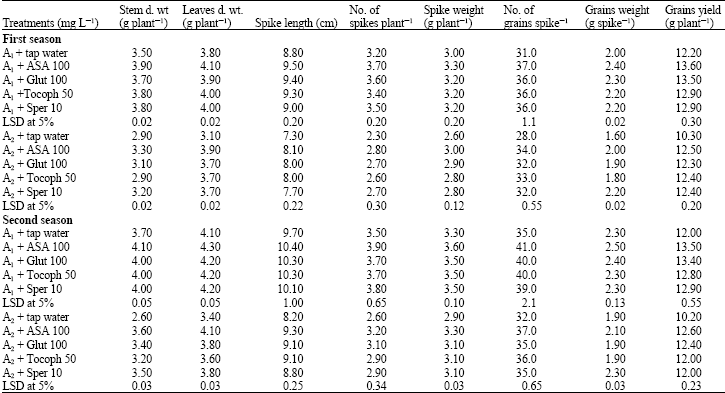 | |
| ASA: Ascorbic acid, Glut.: Glutathione, Tocoph: Tocopherol, Sper.: Spermine, A1: Area 1 (Salinity 3840 mg L-1 or dS m-1), A2: Area 2 (Salinity 6080 mg L-1 or 9.5 dS m-1) | |
| Table 2: | Effect of exogenous applied antioxidants on endogenous enzymatic and non-enzymatic antioxidants in wheat plant grown under soil salt stress conditions (3840 and 6080 mg L-1) during the growing seasons (2006/2007) |
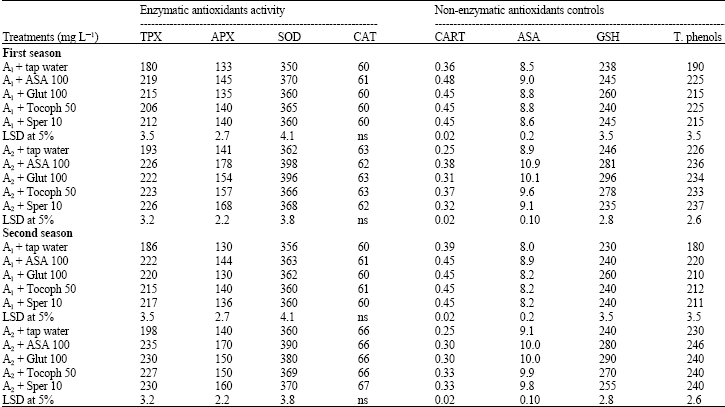 | |
| TPX: Total peroxidase activity (units/g fresh weight), APX: Ascorbic peroxidase activity (units/g fresh weight), SOD: Superoxide dismutase activity (units/mg protein/min), CAT: Catalase activity μmol H2O2 red/mg protein/min), CART: Carotenoids content (mg/g fresh weight), ASA: Ascorbic acid content (mg/g fresh weight), GSH: Reduced glutathione (μmol g-1 fresh weight), T. phenols: Total phenols content (mg g-1 fresh weight), ASA: Ascorbic acid, Glut.: Glutathione, Tocoph.: Tocopherol, Sper.: Spermine, A1: Area 1 (Salinity 3840 mg L-1 or 6 dS m-1), A2: Area 2 (Salinity 6080 mg L-1 or 9.5 dS m-1) | |
The data also show that applied antioxidant materials could alleviate the harmful effect of high soil salt stress levels on yield and its components of wheat plant and ASA more effective in this respect.
Enzymatic antioxidants activity: The data in Table 2 shows that applied antioxidants significantly increased enzymatic activity of total peroxidase (TPX), Ascorbic peroxidase (APX) and Super Oxide Dismutase (SOD) in the two soil salt areas (A1 and A2) compared with untreated plants throughout the two growing seasons. While, catalase activity did not showed any significant response and ASA was the most effective in this respect. It could be noticed that enzymatic activity in A2 was more than that in A1 throughout the growing seasons.
Non enzymatic antioxidant contents: The data in Table 2 shows that any of applied antioxidants increased the contents of endogenous non enzymatic antioxidants (carotenoids, ascorbic acid, glutathione and total phenols). Ascorbic acid treatment proved to be more effective in improving endogenous non-enzymatic antioxidants and the increasing effect in A1 was higher than that of A2 throughout the two growing seasons.
DISCUSSION
The inhibitory effect of soil salt stress areas (A1 and A2) on wheat growth in the present investigation may be due to a decrease in water absorption, metabolic processes, merestematic activity and/or cell enlargement (Sakr et al., 2007) or by damaging growth cells so that they can not perform their functions (Chen and Murata, 2002).
The reduction in seed yield caused by soil salt stress in the two areas (A1 and A2) is largely due to (1) reduction in pollen viability which has been related to decreased calcium mobilization from plant leaves treated with sodium chloride, which is important in pollen germination and pollen tube growth, (2) abscission of flowers or young fruit due to ethylene induction by salinity, (3) moreover, decreasing production of pollen grain, mean number of perfect flowers and fruit set and (4) decreasing the leaf area and number per plant, resulting reduction in the supply of carbon assimilate due to decreasing the net photosynthetic rate and biomass accumulation (Sakr et al., 2007).
As for α-Tocopherol, ascorbic acid and glutathione: It could be concluded that, these plant antioxidants can alleviate the harmful effect of reactive oxygen species (ROS) caused by soil salt stress (3840 and 6080 mg L-1) through several ways such as: (1) inhibiting the lipid photoperoxidation (Thomas et al., 1992), (2) involving in both electron transport of PS II and antioxidizing system of chloroplasts, (3) as membrane stabilizers and multifaceted antioxidants, that scavenge oxygen free radicals, lipid peroxy radicals and singlet oxygen (Diplock et al., 1989), (4) reacting with peroxyl radicals formed in the bilayer as they diffuse to the aqueous phase, (5) scavenging cytotoxic H2O2 and reacts non-enzymatically with other ROS: singlet oxygen, superoxide radical and hydroxyl radical (Blokhina et al., 2003), (6) regenerating another powerful water-soluble antioxidant, ascorbic acid, via the ascorbate-glutathione cycle, (7) stabilize membrane structures (Blokhina, 2003), (8) modulating membrane fluidity in a similar manner to cholesterol and also membrane permeability to small ions and molecules (Foyer, 1992) and (9) decreasing the permeability of digalactosyldiacyl glycerol vesicles for glucose and protons (Berglund et al., 1999).
Regarding polyamines (PAs): It has been suggested that PAs may play a role in antioxidative system and protect membrane from peroxidation. The alleviating effect of polyamines on plants grown under salinity stress may be due to one or more of the following factors: (1) through activating antioxidative defense system, (2) suppressed the level of superoxide and H2O2 in leaf stressed plants (Hernandez et al., 1995), (3) Suppress H2O2 level and thereby membrane damage is being evaluated in terms of antioxidative system, (4) caused reduction in ROS through quenching of singlet oxygen and excited chlorophyll by elevating level of CAR thereby maintained chloroplastic membrane (Velikova et al., 2000), (5) reduce membrane leakage and lipid peroxidation and decreased MDA contents in sugarcane leaves (Zhang and Kirkham, 1996), (6) stabilization of membrane damage may be due to its polycationic nature (Tiburcio et al., 1994), (7) increasing AXP and GR activity as well as CAR and GSH at all salinity levels (Tiburcio et al., 1994), (8) stimulation of chlorophyll synthesis and prevent chlorophyll degradation (Krishnamurthy, 1991) and (9) increasing all organic concentrations, that may be attributed to that polyamines are involved in important biological processes, e.g., ionic balance and DNA, RNA and protein synthesis.
Generally speaking, the application of antioxidants proved to be more effective in alleviating the harmful effect of salinity on wheat plant.
Regarding enzymatic antioxidant activity: Dash and Panda (2001) reported that higher activity of antioxidant enzymes viz., SOD, GR and CAT caused lower H2O2 production, lipid peroxidation and higher membrane stability. Beneficial effect of higher osmolyte concentration (soluble sugars, glycine-betaine, proline and potassium) is reflected in stabilization of essential enzyme proteins such as SOD and GR resulting in higher activity under salinity stress.
Plants posses antioxidant systems in the form of enzymes such as SOD, APX, GR, DIIAR and metabolites viz., ascorbic acid, glutathione, α-tocopherol, carotenoid and flavonoids etc. These antioxidant enzymes and metabolites are reported to increase under various environmental stress as well as comparatively higher activity has been reported in tolerant cultivars than the susceptible ones (Sairam and Srivastava, 2002).
Non enzymatic antioxidants: Ascorbic acid has been implicated in several types of biological activities in plants: (1) as an enzyme co-factor, (2) as an antioxidant and (3) as a donor/acceptor in electron transport at the plasma membrane or in the chloroplasts, all of which are related to oxidative stress resistance (Conklin, 2001).
Total phenols in the wheat leaves tended to increase gradually with increasing salinity levels in soil as shown in Table 2. This increase showed some tendency of wheat to adjust osmotically against salt stress. Moreover, stress condition leads to an increase in phenolic compounds (Naimiki, 1990). These phenolic compounds could be a cellular adaptive mechanism for scavenging oxygen free radicals during stress and this free radical scavenger and others such as ascorbate could be readily oxidized in the system of tissue representing sub-cellular damages.
Reducing glutathione (GSH) can act as an antioxidant in many ways. (1) It can react chemically with singlet oxygen, superoxide and hydroxyl radicals and therefore function directly as a free radical scavenger (Noctor and Foyer, 1998), (2) GSH may stabilize membrane structure by removing acyl peroxides formed by lipid peroxidation reactions and (3) GSH is the reducing agent that recycles ascorbic acid from its oxidized to its reduced form by the enzyme dehydroascorbate reductase (Loewus, 1988).
The increase in reducing glutathione (GSH) contents as reported in Table 2 may be due to the role of the enzymes ascorbate peroxidase, glutathione reductase and superoxide dismutase which involved in the regeneration of glutathione and ascorbate that are important in detoxification of ROS (Foyer et al., 1994).
Generally, it could be concluded that α-tocopherol, ascorbic acid and glutathione can help to alleviate the harmful effect of ROS may be through several ways such as: (1) inhibits the lipid photoperoxidation (Thomas et al., 1992), (2) involves in both electron transport of PS II and antioxidizing system of chloroplasts (Thomas et al., 1992), (3) membrane stabilizers (Thomas et al., 1992), (4) can react with peroxyl radicals (Sairam and Servastava, 2002), (5) it scavenges cytotoxic H2O2 and reacts non-enzymatically with other ROS (Sairam and Servastava, 2002), (6) regenerates another powerful water-soluble antioxidant and ascorbic acid (Blokhina et al., 2002), (7) stabilizes membrane structures (Blokhina, 2002), (8) modulates membrane fluidity in a similar manner to cholesterol and also membrane permeability to small ions and molecules (Foyer, 1992) and (9) decrease the permeability of digalactosyldiacylglycerol vesicles for glucose and protons (Berglund et al., 1999).
As for, role of exogenous antioxidants polyamine on alleviating salinity stress effects: the polyamines (PAs) including spermine (SPM, a tetramine), spermidine (SPD, a triamine) and their obligate precursor putrescine (a diamine) are implicated in induction of plant adaptation to stress (Mishra et al., 2003). It has been suggested that PAs may play role in antioxidative system and protect membrane from peroxidation. The alleviating effect of polyamines on plants grown under salinity stress was previously discussed.
From the above mentioned results it could be noticed that the applied antioxidants could alleviate or minimize the harmful effect of NaCl salinity on wheat plant growth and ascorbic acid and spermine proved to be more effective in this respect.
REFERENCES
- Amako, K., G.X. Chen and K. Asada, 1994. Separate assays specific for ascorbate peroxidase and guaiacol peroxidase and for the chloroplastic and cytosolic isozymes of ascorbate peroxidase in plants. Plant Cell Physiol., 35: 497-504.
Direct Link - Berglund, A.H., R. Nilsson and C. Liljenberg, 1999. Permeability of large unilamellar digalactosyldiacylglycerol vesicles for protons and glucose-influence of α-tocopherol, �-carotene, zeaxanthin and cholesterol. Plant Physiol. Biochem., 37: 179-186.
CrossRefDirect Link - Bettany, A.J.E., 1995. Stress responses in cell cultures of Lolium temulentum. I. Transcriptional and post-transcriptional changes in gene expression during heat shock and recovery. J. Plant Physiol., 146: 162-168.
CrossRefDirect Link - Blokhina, O., E. Virolainen and K.V. Fagerstedt, 2003. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot., 91: 179-194.
CrossRefPubMedDirect Link - Chen, T.H.H. and N. Murata, 2002. Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes. Curr. Opin. Plant Biol., 5: 250-257.
CrossRefPubMedDirect Link - Conklin, P., 2001. Recent advances in the role and biosynthesis of ascorbic acid in plants. Plant Cell Environ., 24: 383-394.
PubMed - Diaz, D.H. and G.C. Martin, 1972. Peach seed dormancy in relation to endogenous inhibitors and applied growth substances. J. Am. Soc. Hortic. Sci., 97: 652-654.
Direct Link - Dash, M. and S.K. Panda, 2001. Salt stress induced changes in growth and enzyme activities in germinating Phaseolus mungo seeds. Biologia Plantarum, 44: 587-589.
CrossRefDirect Link - De Vos, C.H., M.J. Vonk, R. Vooijs and H. Schat, 1992. Glutathione depletion due to copper-induced phytochelatin synthesis causes oxidative stress in Silene cucubalus. Plant Physiol., 98: 853-858.
Direct Link - Dhindsa, R.S., P. Plumb-Dhindsa and T.A. Thorpe, 1981. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation and decreased levels of superoxide dismutase and catalase. J. Exp. Bot., 32: 93-101.
CrossRefDirect Link - Fielding, J.L. and J.L. Hall, 1978. A biolchemical and cytochemical study of peroxidase activity in roots of Pisum sativum I. A comparison of dab-peroxidase and guaiacol-peroxidase with particular emphasis on the properties of cell wall activity. J. Exp. Bot., 29: 969-981.
CrossRefDirect Link - Foyer, M.J., 1992. The antioxidant effects of thylakoid Vitamin E (α-tocopherol). Plant Cell Environ., 15: 381-392.
CrossRefDirect Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hernandez, J.A., E. Olmos, F.J. Corpas, F. Sevilla and L.A. del Rio, 1995. Salt-induced oxidative stress in chloroplasts of pea plants. Plant Sci., 105: 151-167.
CrossRefDirect Link - Krishnamurthy, R., 1991. Amelioration of salinity effect in salt tolerant rice (Oryza sativa L.) by foliar application of putrescine. Plant Cell Physiol., 35: 699-703.
Direct Link - Namiki, M., 1990. Antioxidants/antimutagens in food. Crit. Rev. Food Sci. Nutr., 29: 273-300.
CrossRefPubMedDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Omaye, S.T., J.D. Turnbull and H.E. Sauberlich, 1979. Selected methods for the determination of ascorbic acid in animal cells, tissues and fluids. Methods Enzymol., 62: 3-11.
CrossRefPubMedDirect Link - Sairam, R.K. and G.C. Srivastava, 2002. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci., 162: 897-904.
CrossRefDirect Link - Shalata, A. and P.M. Neumann, 2001. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot., 52: 2207-2211.
CrossRefDirect Link - Thomas, C.E., L.R. McLean, R.A. Parker and D.F. Ohlweiler, 1992. Ascorbate and phenolic antioxidant interactions in prevention of liposomal oxidation. Lipids, 27: 543-550.
CrossRefDirect Link - Tiburcio, A.F., T. Besford, A. Capell, P.S. Borrell, S. Testillano and S. Risueno, 1994. Mechanisms of polyamine action during senescence responses induced by osmotic stress. J. Exp. Bot., 45: 1789-1800.
CrossRefDirect Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link - Velikova, V., I. Yordanov and A. Edreva, 2000. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci., 151: 59-66.
CrossRefDirect Link - Vierling, E., 1991. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 579-620.
CrossRefDirect Link - Zhang, J. and M.B. Kirkham, 1996. Lipid peroxidation in sorghum and sunflower seedlings as affected by ascorbic acid, benzoic acid and propyl gallate. J. Plant Physiol., 149: 489-493.
CrossRefDirect Link - Zhao, F.G. and P. Qin, 2004. Protective effect of exogenous polyamines on root tonoplast function against salt stress in barley seedlings. Plant Growth Regul., 1: 97-103.
Direct Link