
Md. Ariful Islam
College of Veterinary Medicine, Chonbuk National University, Jeonju 561-756, Republic of Korea
Mst. Minara Khatun
College of Veterinary Medicine, Chonbuk National University, Jeonju 561-756, Republic of Korea
Byeong Kirl Baek
College of Veterinary Medicine, Chonbuk National University, Jeonju 561-756, Republic of Korea
Sung Il Lee
Division of Model Animal, Institute of Biomedical Science, Kansai Medical University, Fumizono 10-15, Moriguchi, Osaka, 570-8506, Japan
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 4 | Page No.: 353-359







ABSTRACT
The aim of this study was to assess the impact of Brucella abortus biotype 1 infection on the reproductive performance using Sprague-Dawley (SD) rat model. Virgin female SD rats (n = 24) were infected intraperitoneally with 0.1 mL of saline containing 1x1011 Colony Forming Unit (CFU) of pathogenic B. abortus biotype 1 Korean bovine isolate. Control rats (n = 24) were inoculated with 0.1 mL of apyrogenic saline. Both inoculated and control rats were divided into six subgroups. Four rats in each subgroup were consistently bred at 3, 7, 14, 21, 28 and 60 days after infection. B. abortus infection induced 41.67% infertility in the infected rats. The mean number of offspring/litter was 8.71±2.01 for infected rats and 12.87±1.42 for control rats (p<0.001). The mean weight of the viable offspring was 6.10±0.36 g for infected rats and 7.15±0.40 g for control rats (p<0.001). The rate of stillbirth was 12.30% in the infected rats. B. abortus biotype 1 was isolated from the uteri of the infected rats. The data of this study indicate that B. abortus biotype 1 infections in SD rat model affect reproduction adversely by causing infertility, stillbirth and loss of number and weight of offspring.
PDF Abstract XML References Citation
How to cite this article
Md. Ariful Islam, Mst. Minara Khatun, Byeong Kirl Baek and Sung Il Lee, 2009. Effects of Brucella abortus Biotype 1 Infection on the Reproductive Performance of Sprague-Dawley Rats. Pakistan Journal of Biological Sciences, 12: 353-359.
DOI: 10.3923/pjbs.2009.353.359
URL: https://scialert.net/abstract/?doi=pjbs.2009.353.359
DOI: 10.3923/pjbs.2009.353.359
URL: https://scialert.net/abstract/?doi=pjbs.2009.353.359
INTRODUCTION
Brucellosis is an important zoonosis, caused by several species of the genus Brucella that affect numerous domestic and wild mammals and humans (Whatmore et al., 2006; Macedo et al., 2008). Brucella abortus is a potentially pathogenic bacterium for the reproductive tract of cattle (Givens, 2006) that can be introduced into the uterus during natural breeding with the infected male (Crawford et al., 1990) and artificial insemination with contaminated semen (Lim et al., 2005). Localization of brucellae within reproductive tracts accounts for the principle clinical symptoms of abortion and/or reduced fertility in most natural host species (Robison et al., 1998). The major clinical manifestation of brucellosis in cattle and wildlife is abortion, decreased fertility and retention of placenta (Enright, 1990; Yaeger and Holler, 1997; Rhyan et al., 1994). Uterine discharge of aborted fetus is believed to be principle source of infection to other animals. Brucellosis in humans resulted from direct contact with the infected animals as well as consumption of contaminated milk and milk products (Pappas et al., 2006). Undulant fever, arthritis and endocarditis are the clinical manifestations in human brucellosis (Young, 1995). Brucellosis accounts huge economic losses in livestock industries due to loss of offspring, increased calving interval, decreased fertility and culling of the infected animals (Radostits et al., 2007).
Brucellosis has emerged as a serious animal and public health issue in many developing countries (Pappas et al., 2005) including Korea (Park et al., 2005) despite animal control and eradication program. Brucella infection in both cattle and people in Korea is commonly due to B. abortus biotype 1 (Park et al., 2005; Wee et al., 2008).
Rat model was used for the evaluation of the vertical transmission of B. abortus biotype 1 (Baek et al., 2005). In the present study we investigated the effects of B. abortus biotype 1 infection on the reproductive performance in SD rat model with regards to the inoculation period. We infected sexually mature virgin female SD rats with B. abortus biotype 1 bovine pathogenic Korean isolate and noted infertility, stillbirth, losses of weight and number of offspring among infected rats after breeding.
MATERIALS AND METHODS
Ethics: The experimental protocols were approved by the Animal Care and Use Committee at the Chonbuk National University and were in accordance with the Chonbuk National University guide for the Care and Use of Laboratory Animals.
Experimental rats: Ten to twelve-week-old SD male (n = 12) and female virgin rats (n = 48) weighing 250-300 g were used in this study. All of these rats were obtained from a Specific Pathogen Free (SPF) laboratory animal company (Koatech, Pyungtaek city, Gyeonggido 451-864, Korea). The rats were housed in a stringently hygienic, climate-controlled environment and provided with commercial food (Koatech, Pyungtaek city, Gyeonggido 451-864, Korea) and water ad libitum. The animals were culture negative for Brucella infection and seronegative for B. abortus antibodies, prior to the experimental infection, ascertained by routine bacteriological and serological tests.
Bacterial strain and media: Pathogenic B. abortus biotype 1 originating from cattle in Korea was used for experimental infection in rats. The bacteria were maintained as frozen glycerol stocks and cultured on brucella agar media (Difco, Kansas City, Missouri, USA) for 5-7 days at 37°C with 5% CO2. The grown bacteria were harvested in normal saline.
Experimental inoculations: The female rats were divided into an infected group (n = 24) and a control group (n = 24). All rats in infected group were injected intraperitoneally with 0.1 mL of normal saline containing 1x1011 CFU mL-1 of the B. abortus biotype 1. All control rats were injected intraperitoneally with 0.1 mL apyrogenic saline.
Clinical examinations: Clinical parameters, such as body temperature, appetite and thirst of the infected and control rats were recorded in the morning for a period of 7 days.
Breeding plan: To assess the effect of infection on the reproductive performance in relation to the inoculation period, the infected rats (n = 24) were divided into six subgroups for breeding. Four rats in each subgroup were separately bred at 3, 7, 14, 21, 28 and 60 days after infection with one uninfected healthy male rat (1 male for 4 females). Control rats (n = 24) were also divided into six subgroups and bred simultaneously with infected subgroups following the similar breeding protocol of infected rats. One male rat was kept with four female rats in a single cage for a period of one month. Pregnant rats were identified by abdominal distension and placed in individual cages 3 or 4 days before parturition. The pregnant rats were continuously monitored and after parturition number of live or stillborn offspring were counted and weight of the live offspring were measured.
Reproductive performance: Reproductive performance in infected and control rats were assessed by counting or measuring the number of live or stillborn offspring/dam and weight of day-old live offspring.
Serological studies: After parturition, 3 mL of blood samples were collected from each of the infected rats by aseptic cardiac puncture under general anesthesia induced by intraperitoneal administration of 7-25 mg kg-1 of Tiletamine and Zolazepam (Zoletil 50®, Virbac Laboratories-06515 Carros-France). Blood samples were also collected from the non-pregnant infected female rats as well as male rats (n = 6) used for breeding with the infected female rats. Serum samples were collected and stored in small aliquots at -20°C until tested. Serum samples were screened for anti-Brucella antibodies by the Rose Bengal Plate Agglutination Test (RBPAT) as described previously (Alton et al., 1988). Sera of the parturient infected rats bred at 3, 7, 14, 21, 28 and 60 days after infection were examined in the enzyme-linked immunosorbent assay (ELISA) against Brucella smooth-lipopolysaccharide (S-LPS) by using the anti-IgG1 and anti-IgG2a conjugates. The ELISA was performed as described previously (Lamb et al., 1979) with little modification. Flat-bottomed 96-well polysterine microtiterplates (Nunc, Denmark) were coated with 100 μL of LPS (5 μg mL-1) of B. abortus strain 1119-3 suspended in 0.05 mM sodium bicarbonate buffer (pH 9.6). Affinity purified rat IgG1 (Bethyl Laboratories, Inc, USA) and IgG2a (Bethyl Laboratories, Inc, USA) were used to coat the 96-well plate starting from 500 to 7.8 ng well-1 for generation of standard curve. Each plate was incubated at 4°C overnight. Plates were washed three times with wash solution (PBST: PBS (pH 7.4) with 0.05% (v/v) Tween 20). Each well of the antigen-coated plates were blocked with 200 μL of blocking solution of 1% (w/v) bovine serum albumin (Sigma Aldrich Inc., St. Louis, MO, USA) in PBST and incubated at 37°C for 30 min. After three washes with PBST, 100 μL of sera samples, diluted 1:100 in sample diluent (50 mM tris, 0.14 M NaCl, 1% BSA, 0.05% Tween 20, pH 8.0) were added to each well in duplicate. The plates were sealed and incubated at 37°C for 1 h. After five washing cycles with PBST, each well was incubated with 100 μL of 1:100000 dilution of goat anti-rat IgG1 and IgG2a antibodies conjugated to horseradish peroxidase (Bethyl Laboratories Inc, USA) diluted in conjugate diluent (50 mM tris, 0.14 M NaCl, 1% BSA, 0.05% Tween 20, pH 8.0) and the plates incubated at 37°C for 1 h. After five washings as described above, the color reaction was developed by adding 200 μL well-1 of a solution containing 1.0 mg mL-1 of O-phenylenediamine dihydrochloride (OPD; Sigma, St. Louis, USA) in 0.05 M citrate buffer (pH 4.0) with 0.04% (v/v) H2O2. The plates were incubated in the dark for 30 min at room temperature. The colorimetric reaction was stopped by the addition of 50 μL well-1 of 3 M H2SO4. The absorbance measurements were made at 492 nm, using an automatic ELISA plate reader.
Bacteriological studies: After parturition all infected and live offspring born from them were euthanized. Non-pregnant infected as well as male rats (n = 6) used for breeding with infected females were also euthanized. Specimens of uterus and spleen were collected from all of the infected rats. Liver and spleen were collected from all of the live and stillborn offspring. Spleen and testes were collected from the six male rats. All samples were stored at -20°C until cultured. Thawed tissues were macerated in a stomacher (Costa Brava, Spain). All macerated samples were plated on brucella agar media (Difco, Kansas City, Missouri, USA) supplemented with antibiotics (cycloheximide, polymixin B and bacitracin that inhibit growth of bacteria other than Brucella) and incubated at 37°C with 5% CO2 for 5-7 days. The identification of the isolates in the culture positive specimens was conducted by routine methods (Alton et al., 1988).
Polymerase chain reaction: Bacteria harvested from culture positive specimens of rats were also confirmed to be B. abortus biotype 1 by AMOS (abortus, melitensis, ovis, suis) Polymerase Chain Reaction (PCR) described previously (Bricker and Halling, 1994). For AMOS-PCR profiling, DNA was extracted from Brucella suspected colonies by a genomic DNA extraction kit (AccuPrep DNA Extraction Kit, Bioneer, Daejeon, Korea) using manufacture’s protocol.
Histopathological analysis: Uteri collected from infected and control pregnant rats were fixed in 10% neutral buffered formalin. The fixed tissues were processed, sectioned and stained with Hematoxylin-Eosine (HE) and examined for histopathological changes under a light microscope (Meador et al., 1988).
Statistical analysis: All statistical analysis was conducted by the Student’s t-test. Significance of all the analyses was established at p-value of <0.05.
RESULTS
Clinical findings: All rats infected with B. abortus biotype 1 became lethargic, anorexic and febrile within 24 h. The highest rectal temperature was 38.5°C at 72 h within 7 days observation period. No abnormal clinical signs were observed in control rats through out the experiment.
Reproductive performance: In six infected subgroups 25 to 75% rats became pregnant and the mean litter size and body weight of day-old offspring were ranged from 6.33±0.57 to 11.5±3.53 and 5.7±0.28 to 6.6±0.31 g, respectively. Stillbirth was recorded in all of the infected subgroups. Among six infected subgroups the lowest rate of pregnancy (25%) and highest rate of stillbirth (40%) were recorded in rats bred at 14 days after infection. In all control subgroups 100% rats became pregnant and the mean litter size and body weight of day-old offspring were ranged from 12±1.63 to 13.5±1.29 and 6.92±0.51 to 7.30±0.58 g, respectively. The reproductive performance of infected and control rats in six subgroups are shown in Table 1.
| Table 1: | Reproductive performance of the B. abortus biotype 1 infected and control rats bred at 3, 7, 14, 21, 28 and 60 days after infection |
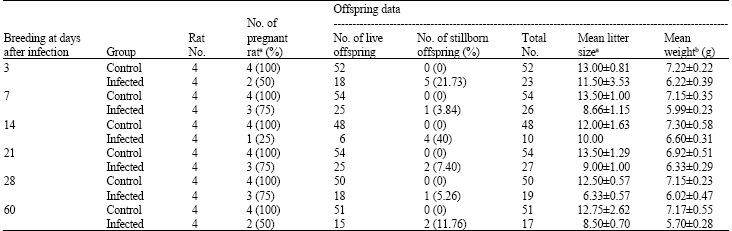 | |
| aRat that delivered offspring, bDay-old live offspring only | |
| Table 2: | Summary of the reproductive performance of B. abortus biotype 1 infected and control rats |
 | |
| aLive and stillborn offspring only, bNot determined | |
| Table 3: | Results of isolation and identification of B. abortus biotype 1 from infected female rats and offspring born to them by the routine methods and AMOS-PCR assay |
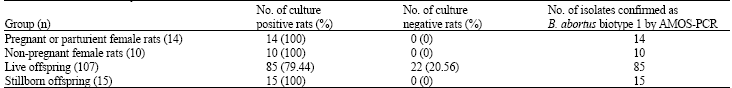 | |
A total of 14 rats out of 24 infected rats became pregnant and delivered 122 offspring. Out of 122 offspring, 107 were live and 15 were stillborn. All control rats were pregnant and delivered 309 live offspring. The overall reproductive performance of infected as well as control rats is summarized in Table 2.
Bacteriological findings: Characteristic colonies of B. abortus biotype 1 were cultured from uteri and spleen of all infected female rats. Out of 107 live offspring, 85 offspring born to infected dams were culture positive for B. abortus biotype 1. On the other hand, B. abortus biotype 1 was isolated from all of the stillborn offspring born to infected dam. No bacteria were isolated from any of the six male rats bred with infected females. The results of isolation and identification of B. abortus biotype 1 cultured from specimens of infected female rats and offspring born to them is summarized in Table 3.
Polymerase chain reaction: All culture positive bacterial colonies harvested from specimens of infected female rats as well as offspring born to them were confirmed as B. abortus biotype 1 by AMOS-PCR with the predicted 498-base pair PCR amplicons (Fig. 1).
Serological findings: Serum samples of the infected pregnant and non-pregnant rats were tested positive in the RBPAT screening test. Serum samples of breeding male rats were tested negative by RBPAT screening test. We determined the magnitude and Th-specificity of antibodies in sera of the infected pregnant rats generated against S-LPS of B. abortus by the indirect ELISA. In the sera of pregnant rats IgG2a indicative of Th1 response was significantly higher as compare to IgG1 indicative of Th2 response (p<0.001). The results of ELISA are presented in Fig. 2.
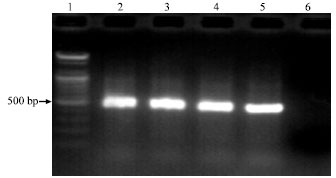 | |
| Fig. 1: | AMOS-PCR profile for amplification of DNA of B. abortus. Lanes: (1), 100 bp size DNA marker (Bioneer, Daejeon, Korea), (2) bacterial DNA of an infected female rat, (3) bacterial DNA of a stillborn offspring, (4) bacterial DNA of a live offspring, (5) positive control with DNA of B. abortus strain 1119-3, (6) negative control without DNA |
 | |
| Fig. 2: | Serum IgG1 and IgG2a responses measured by ELISA in B. abortus biotype 1 infected pregnant rats bred at 3, 7, 14, 21, 28 and 60 days after infection. The responses are presented as Mean±SD. Statistically significant difference between IgG1 and IgG2a are indicated by asterisks (**p<0.001) |
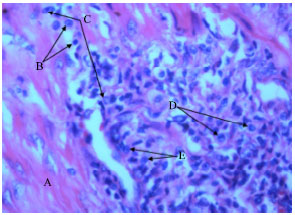 | |
| Fig. 3: | Histopathological findings. The lesions in the endometrium of B. abortus infected uterus of a pregnant rat characterized by A: Muscular thickening, B: Accumulation of lymphocytes, C: Neutrophils, D: Monocytes and E: Macrophages, magnification 400x |
Histopathological findings: Histopathological analysis of the uteri of the infected pregnant rats, compare to analysis of the control rat uteri, showed some lesions in the endometrium. Histopathological lesions of the endometrium of infected pregnant rats were characterized by thickened muscular wall with the moderate to diffuse infiltration of inflammatory cells such as lymphocytes, neutrophils, monocytes and macrophages (Fig. 3).
DISCUSSION
In the present study we investigated the effects of B. abortus biotype 1 infection on the reproductive performance in the SD rat model. We noted infertility, stillbirth and loss of number and weight of the offspring among infected rats in this study. These data are discussed in relation to the bacteriological, serological and pathological findings.
In this study, we compare the reproductive performance of the infected rats with the control rats to figure out the effect of B. abortus biotype 1 infection on the reproductivity. We included control rats in all subgroups and simultaneously bred with the infected rats in order to match age during the time of breeding. In the present study, the highest infertility rate was seen in the infected rats bred at 14 days after infection (Table 1). The overall 41.67% infertility was recorded in the infected rats (Table 2) which were resulted from reproductive tract infection since B. abortus biotype 1 was isolated from the nongravid uteri of all of the infected female rats (Table 3). All of the non-pregnant infected rats elicited antibody responses against B. abortus biotype 1 in the RBPAT screening test also support the establishment of infection. Uterine infection is a key risk factor for abnormal ovarian hormone profile and subfertility affecting up to 44% of animals (Royal et al., 2000). Bacterial infection also affects the function of uterus. Endometrial and ovarian cell functions were modified by challenge with bacteria, their product such as LPS or proinflammatory cytokines (Herath et al., 2006). LPS is the component of the cell wall of B. abortus that act as a virulent factor (Lapaque, 2005).
Infection causes damage to the uterine tissue and histological lesions of endometrium in the animal are associated with the poor reproductive performance (Bonnett, 1993). In this study, B. abortus biotype 1 was isolated from all of the pregnant rats after parturition which caused lesions of the uteri (Fig. 3). In cattle, the most destructive lesions of brucellosis occur in the gravid uterus consisted of changes in the interstitial tissue and accumulations of lymphocytes, macrophages and neutrophils (Deyoe, 1980).
Infectious agents can be responsible for reduced ovulation rates, fertilization rates and embryonic survival rates (Givens, 2006). Uterine infection precludes fertilization or embryo survival (Semambo et al., 1991). In the present study, the mean number of offspring per litter was significantly lower in infected rats (8.71±2.01) compare to that of control rats (12.87±1.42) (p<0.001) (Table 2). Among infected subgroups the mean litter size was lower in rats bred at 28 days after infection (Table 1).
In mice, B. abortus colonization of the gravid reproductive tract leads to placental damage, fetal infection and fetal dead (Bosseray, 1982; Tobias et al., 1993). In the present study, 12.30% of the offspring born to infected rats were stillborn which were found to be infected with B. abortus biotype 1 (Table 3). Stillbirths were reported in cattle infected with B. abortus (Kirkbride, 1993). The highest rate of stillbirth was recorded in rats bred at 14 days after infection (Table 1).
The mean weight of the day-old live offspring (6.10±0.36 g) born to the infected rats was significantly lower as compared to the offspring born to the control rats (7.15±0.40 g) (p<0.001) (Table 2). The lowest mean body weight of offspring was recorded in rats bred at 60 days after infection (Table 1). In Bartonella birtless infection, the weight of viable fetuses was significantly lower which were resulted from fetal suffering during gestation (Boulouis et al., 2001). In this study, loss of weight of live offspring born to B. abortus infected rats might have resulted from fetal suffering during gestation.
There was no abortion recorded in any of the infected rats in our study. In cattle when they are infected before pregnancy seldom abort during subsequent pregnancy (Deyoe, 1980).
There is a relationship between immune response and outcome of pregnancy. In Brucella infections, cellular immune response plays a predominant role compared with the humoral response (Golding et al., 2001). Cellular immune response against B. abortus infection is primarily mediated by Th1 type of immune response which adversely affect pregnancy outcome (Krishnan et al., 1996). In this study, among infected pregnant rats Th1 type of immune response was significantly dominant (p<0.001) as compared to Th2 response against B. abortus biotype 1 infection (Fig. 2). Cellular immune response against B. abortus infection might affect the reproductive performance of the infected pregnant rats in this study.
Present study demonstrated that infection of SD rats with the B. abortus biotype 1 Korean bovine isolate resulted low fertility, stillbirth and loss of number and weight of the offspring. Intervention measures against reproductive tract infection caused by B. abortus need to be addressed.
REFERENCES
- Alton, G.G., L.M. Jones, R.D. Angus and J.M. Verger, 1988. Techniques for the Brucellosis Laboratory. 1st Edn., Institute Nationale de le Rech, France, Paris, Pages: 174.
Direct Link - Baek, B.K., B.O. Lee, J. Hur, M.S. Rahman, S.I. Lee and I. Kakoma, 2005. Evaluation of the Sprague-Dawley rat as a model for vertical transmission of Brucella abortus. Can. J. Vet. Res., 69: 305-308.
PubMedDirect Link - Bonnett, B.N., S.W. Martin and A.H. Meek, 1993. Associations of clinical findings, bacteriological and histological results of endometrial biopsy with reproductive performance of postpartum dairy cows. Prev. Vet. Med., 15: 205-220.
Direct Link - Boulouis, H.J., F. Barrat, D. Bermond, F. Bernex, D. Thibault, R. Heller, J.J. Fontaine, Y. Yves Piemont and B.B. Chomel, 2001. Kinetics of Bartonella birtlesii infection in experimentally infected mice and pathogenic effect on reproductive functions. Infect. Immun., 69: 5313-5317.
PubMed - Bricker, B.J. and S.M. Halling, 1994. Differentiation of Brucella abortus bv. 1, 2 and 4, Brucella melitensis, Brucella ovis and Brucella suis bv. 1 by PCR. J. Clin. Microbiol., 32: 2660-2666.
PubMedDirect Link - Givens, M.D., 2006. A clinical, evidence-based approach to infectious causes of infertility in beef cattle. Theriogenology, 66: 648-654.
CrossRef - Golding, B., D.E. Scott, O. Scharf, L.Y. Huang and Y. Zaitseva et al., 2001. Immunity and protection against Brucella abortus. Microbes Infect., 3: 43-48.
CrossRefPubMedDirect Link - Krishnan, L., L.J. Guilbert, T.G. Wegmann, M. Belosevic and T.R. Mosmann, 1996. T Helper 1 response against Leishmania major in pregnant C57BL/6 mice increases implantation failure and fetal resorptions. Correlation with increased IFN-gamma and TNF and reduced IL-10 production by placental cells. J. Immunol., 156: 653-662.
- Lapaque, N., I. Moriyon, E. Moreno and J.P. Gorve, 2005. Brucella lipopolysaccharide acts as a virulent factor. Curr. Opin. Microbiol., 8: 60-66.
PubMed - Lim, H.S., Y.S. Min and H.S. Lee, 2005. Investigation of a series of brucellosis cases in Gyeongsangbuk-do during 2003-2004. J. Prev. Med. Public Health, 38: 482-488.
PubMed - Macedo, G.C., D.M. Magnani, N.B. Carvalho, O.B. Romero, R.T. Gazzinelli and S.C. Oliveira, 2008. Central role of MYD88-dependent dendritic cell maturation and proinflammatory cytokine production to control Brucella abortus infection. J. Immunol., 180: 1080-1087.
PubMed - Pappas, G., N. Akritidis, M. Bosilkovski and E. Tsianos, 2005. Brucellosis. N. Engl. J. Med., 352: 2325-2336.
PubMedDirect Link - Pappas, G., P. Papadimitriou, N. Akritidis, L. Christou and E.V. Tsianos, 2006. The new global map of human brucellosis. Lancet Infect. Dis., 6: 91-99.
CrossRefPubMedDirect Link - Park, M.Y., C.S. Lee, Y.S. Choi, S.J. Park, J.S. Lee and H.B. Lee, 2005. A sporadic outbreak of human brucellosis in Korea. J. Korean Med. Sci., 20: 941-946.
PubMed - Royal, M.D., A.Q. Darwash, A.P.F. Flint, R. Webb, J.A. Woolliams and G.E. Lamming, 2000. Declining fertility in dairy cattle: Changes in traditional and endocrine parameters of fertility. Anim. Sci., 70: 487-501.
CrossRefPubMedDirect Link - Tobias, L., D.O. Cordes and G.G. Schurig, 1993. Placental pathology of the pregnant mouse inoculated with Brucella abortus strain 2308. Vet. Pathol., 30: 119-129.
Direct Link - Wee, S.H., N.M. Nam and C.H. Kim, 2008. Emergence of brucellosis in cattle in the Republic of Korea. Vet. Rec., 162: 556-557.
PubMed - Whatmore, A.M., S.J. Shankster, L.L. Perrett, T.J. Murphy, S.D. Brew, R.E. Thirlwall, S.J. Cutler and A.P. MacMillan, 2006. Identification and characterization of variable-number tandem-repeat markers for typing of Brucella spp. J. Clin. Microbiol., 44: 1982-1993.
PubMed - Young, E.J., 1995. An overview of human brucellosis. Clin. Infect. Dis., 21: 283-289.
CrossRefPubMedDirect Link