
Abeer A. Ali
Department of Mycological and Plant Diseases Survivals, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
K.M. Ghoneem
Department of Mycological and Plant Diseases Survivals, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
M.A. El-Metwally
Department of Mycological and Plant Diseases Survivals, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
K.M. Abd El-Hai
Department of Leguminous and Forage Crops Disease, Plant Pathology Research Institute, Agriculture Research Center, Giza, Egypt
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 3 | Page No.: 213-221







ABSTRACT
Root rot caused by soil borne pathogenic fungi is the most sever disease attacks lupine plants. Isolation trials from diseased plants in some areas of Dakahlia Province (Egypt) was carried out. Rhizoctonia solani and Fusarium solani proved to be the most dominant isolates. Meanwhile, Fusarium oxysporum and Sclerotium rolfsii were less frequent. Efficacies of some plant resistance elicitors vis: chitosan (CHI), salicylic acid (SA) and hydroquinone (HQ) in comparing to the fungicide Rhizolex T-50 as seed treatments showed significant reduction in the fungal growth in vitro. Chitosan at 8 g L-1 and fungicide completely inhibited the growth of all isolated fungi, while SA at 1.4 g L-1 and HQ at 1.2 g L-1 inhibited the growth of Fusarium solani and F. oxysporum, respectively. The greenhouse experiments showed that S. rolfesii (No. 6) and R. solani (No. 2) followed by F. solani (No. 5) and F. oxysporum (No. 9) were the most aggressive root rot fungi. Soaking susceptible lupine seeds (Giza 1) in each one of the three selected elicitors showed a significant reduction in seedlings mortality. CHI at 8 g L-1 was superior in increasing the percentage of healthy plants to record 72.5, 80.9, 62.7and 64.3%, when seeds were grown in soil infested with of F. solani, F. oxysporum, R. solani and S. rolfesii, respectively. These results were confirmed under field conditions in two different locations i.e., Tag El-Ezz and El-Serow Research Stations. CHI 8 g L-1 proved to be the best elicitor after fungicide, in reducing lupine root rot disease. It showed 41 and 60% reduction in the plants mortality comparing to 56.37 and 69.13% in case of Rhizolex-T in Tag El-Ezz and El-Serow locations, respectively. The treatments were accompanied with a significant increase in lupine growth parameters, yield components and physiological aspects. Application of CHI at 8 g L-1 or HQ at 1.2 g L-1 was the most potent in this respect as compared to check treatment.
PDF Abstract XML References Citation
How to cite this article
Abeer A. Ali, K.M. Ghoneem, M.A. El-Metwally and K.M. Abd El-Hai, 2009. Induce Systemic Resistance in Lupine Against Root Rot Diseases. Pakistan Journal of Biological Sciences, 12: 213-221.
DOI: 10.3923/pjbs.2009.213.221
URL: https://scialert.net/abstract/?doi=pjbs.2009.213.221
DOI: 10.3923/pjbs.2009.213.221
URL: https://scialert.net/abstract/?doi=pjbs.2009.213.221
INTRODUCTION
The Egyptian lupine (Lupinus termis Forsk) is a fabaceous crop which grown in Egypt for food, medical and industrial purposes. The plant is cultivated for its non-endospermic seeds, which contains alkaloids, protein, oil, cholesterol, lecithin, salts (phosphorus and potassium) and carbohydrates (Ibrahim et al., 1990). The alkaloids especially lupulin is occasionally employ as a stomach tonic and as on hypnotic to promote sleep (Wallis, 1967). Green plants are useful as green-manuring because of the high nitrogenous content (Roberto Chiej, 1984).
Number of soil-borne fungi attack lupine causing decrease in seed yield including Rhizoctonia solani, Sclerotium rolfsii, Fusarium solani and F. oxysporum. They reported to attack lupine roots and stem base; causing serious losses in seed germination and plant stand as well (Fahim et al., 1983; Osman et al., 1986).
Since the control of soil borne pathogens depends mainly on the use, pesticides applications and this increase the input of pesticides into the environment and produce pesticide-resistant of strains pathogen. Efforts are being made to develop alternative protection strategies. One solution might be the activation of the plant’s own defense system, known as induced resistance (Sticher et al., 1997). This induced systemic resistance is triggered by a number of chemicals such as salicylic acid (SA) and hydroquinone (HQ) (El-Mougy et al., 2002; Matraux et al., 1990; Shahda, 2000; Amin et al., 2007) or biologically active oligoglucosides, the so-called elicitors (Benhamou et al., 1994) which have been shown to induce systemic resistance in many crops like tomato, tobacco, pea, maize, cotton, rice, potato and other vegetables against viruses, fungi and bacteria (Oostendorp et al., 2001). Among the most promising bioactive oligosaccharides is chitosan (CHI) which has attracted attention because of its unique biological properties of various pathogenic fungi and ability to be a potent elicitor of plant defense reactions against soil-borne pathogens (Benhamou et al., 1998; Prapagdee et al., 2007).
This research aims to develop and evaluate new or existing alternative safe control method for lupine root rot under greenhouse and field conditions. The research also was planed to study the effect of these treatment on the contents of phenolic compounds and photosynthetic pigments in the coming up plants.
MATERIALS AND METHODS
Fungal pathogens: Samples of lupine plants showing root rot disease symptoms were collected from different locations of Dakahlia regions in 2007 growing season. All samples were thoroughly washed in tap water, cut in small pieces (1 cm) and surface sterilized for 2 min in 2% sodium hypochlorite solution, then rinsed several times in sterilized distilled water and dried between a number folds of sterilized filter papers. The surface sterilized samples were plated onto Potato Dextrose Agar (PDA) medium supplemented with streptomycin-sulfate (100 mg mL-1) and incubated at 27°C. After 4-5 days incubation period, the developed fungal colonies were purified by hyphal tip and single spore isolation techniques. The fungi then were identified according to cultural and microscopically characters described by Ellis (1976), Sneh et al. (1992) and Nelson et al. (1983). The number of each isolated fungus was recorded and the frequency of each fungus in each location was recorded.
Pathogenicity tests: Pathogenicity tests were carried out on 20 isolates of F. solani, 17 isolates of F. oxysporum, 22 isolates of R. solani and 9 isolates of S. rolfesii to choose the most aggressive isolates from each fungal species.
Inocula of the isolated pathogens were prepared using sorghum: coarse sand: water (2:1:2 v/v) medium. The ingredients were mixed, bottled and autoclaved for 20 min at 1.5 air pressure. The sterilized medium was inoculated using agar discs obtained from the periphery of 5 days old colony of each of the isolated fungi. The inoculated media were incubated at 28 ± 1 °C for 15 days. Plastic pots (25 cm diameter) filled up with 4 kg autoclaved clay soil and artificially infested with the previously prepared fungi inoculum at the rate of 5% by weight. Pot soil were mixed thoroughly with inoculum then, watered and left for one week to ensure even distribution of the inoculated fungi. Five pots were used as replicates for each treatment as well as check treatment (uninfested soil). Apparently healthy lupine seeds (cv. Giza 1) were surface sterilized in 0.1% sodium hypochloride for 2 min, then washed in sterilized water. Dried lupine seeds were sown in infested soil (5 seed pot-1). After 15 and 30 days from sowing, seedlings were rated for pre- and post-emergence damping-off, respectively.
Effect of tested elicitors on the fungal growth: The effect of the tested elicitors and the fungicide Rhizolex-T 50 W.P. on linear growth of the isolated fungi was carried out. CHI was dissolved in 0.1% acetic acid in distilled water according to Eikemo et al. (2003). The medium PDA was amended with each elicitor just before solidification. The appropriate amount of each elicitor was prepared by mixing it with 50 mL medium in each flask to give concentrations of 6.0 and 8.0 g L-1 of CHI, 0.6 and 1.2 g L-1 of HQ, 0.7 and 1.4 g L-1 of SA and 3 g L-1 of fungicide. Ten milliter of each PDA medium was poured in each Petri dish, then they were inoculated with 5 mm discs of fungal growth and incubated at 25 °C in the dark. Check dishes having no elicitors or fungicide were inoculated at the same way. Five replicate were used per treatment. The fungal growth was measured when the full growth of tested fungi was observed in the check treatment and the average growth diameter was calculated.
Control of soil-borne fungi under greenhouse condition: Controlling of the most aggressive pathogenic fungi of lupine root rot was carried out under greenhouse conditions. Pots of previously prepared as mentioned above were seeded with soaked lupine seeds in each of the three tested elicitors (CHI, SA and HQ) for 2 h. Just before sowing at the rate of 6 and 8, 0.7 and 1.4 and 0.6 and 1.2 g L-1 water, respectively. Six treated seeds pot-1 were sown in infested soil in comparing with seeds of check and seeds treated with Rhizolex-T 50 fungicide. Soil treated with free medium of the fungus was served as check treatment. Percentages of pre- and post-emergence damping-off were assessed at 15 and 40 days from sowing, respectively.
Field experiment: Filed experiment was carried out at two localities i.e., Tag El-Ezz Research Station, Dakahlia Province and El-Serow Research Station, Damietta Province, Egypt during the growing winter season of 2007/2008. Elicitors treatments were used as seed soaking treatment while, Rhizolex-T 50 w.p. was used as seed coating as mentioned above. Treated lupine seeds (cv. Giza 1) were sown in the field at the 10th of November 2007 in both localities and left under natural infection.
A complete randomized block design with three replicates was used in both experiments. The experimental plot contained 5 ridges occupying an area of 10.5 m2 (3x3.5 m). Percentages of root rot incidence at pre- and post-emergence damping-off were recorded at 15 and 60 days from sowing, respectively.
For morphological characters of lupine, samples were recorded after 120 days from planting including plant height, number of branches and number of leaves plant-1. At harvest (165 days from sowing), number of pods, seed number plant-1, weight of seeds plants-1 and weight of 100 seeds were recorded.
Determination of total phenolic compounds: Total phenol were determined after 70 days from sowing in fresh shoot using the Folin-Ciocalteau reagent (Singleton and Rossi, 1965). Samples (2 g) were homogenized in 80% aqueous ethanol at room temperature and centrifuged at 10000 rpm for 15 min, under cooling and the supernatants were saved. The residues were re-extracted twice in 80% ethanol and supernatants were pooled, put into evaporating dishes and evaporated to dryness at room temperature. Residues were dissolved in 5 mL of distilled water. One-hundred microlitres of this extract were diluted to 3 mL with water and 0.5 mL of Folin-Ciocalteau reagent was added. After 3 min, 2 mL of 20% of sodium carbonate was added and the contents were mixed thoroughly. The developed color was photometrically measured at 650 nm, after 60 min using catechol as a standard. The results were expressed as mg catechol/100 g fresh weight material.
Determination of photosynthetic pigments: The blade of the 3rd leaf from plant tip (terminal leaflet) was taken at 70 days to determine photosynthetic pigments (chlorophyll a, b and carotenoids). Photosynthetic pigments were extracted in methanol 90% for 24 h at room temperature while traces of sodium carbonate was added (Robinson and Britz, 2000). The pigments were determined at the wave lengths (452.5, 650 and 665 nm) as described by Mackinney (1941).
Statistical analysis: Obtained data were statistically analyzed by CoStat 3.4 software as the usual technique of analysis of variance (Gomez and Gomez, 1984). The means were compared using Least Significant Difference (LSD) at p≤0.05 as outlined by Duncan (1955).
RESULTS
Root rot fungi: Sixty eight fungal isolates representing four species of three genera were isolated from roots of lupine plants showing root rot symptoms including Fusarium sp. (37 isolate), Rhizoctonia solani (22 isolate) and Sclerotium rolfsii (9 isolate).
Results in Table 1 indicate that the high frequency isolated fungi (F. solani and F. oxysporum) were found in Temi El-Amdeed distract at percentages of (25.3 and 19.2%) followed by Sherbeen distract (22.4 and 16.6%). While, R. solani was isolated from the diseased plants at Belkas distract in a frequency of (38.5%). At Sherbeen distract it was (36.1%). On the other hand, S. rolfsii occurred in Temi El-Amdeed distract at a percentage of (21.1%) followed by (19.2%) at El-Mansoura distract.
Pathogenicity tests: All tested fungi (68 isolates) were pathogenic and causes typical symptoms of pre- and post-emergence damping-off on lupine seedlings. S. rolfesii showed to be the most virulent fungus to cause 77.3% seedlings mortality, followed by R. solani, F. solani and F. oxysporum (67.7, 58.0 and 50.3%, respectively).
Results from the pathogenicity tests on the 30 days old lupine seedlings indicated that F. solani (No. 5), F. oxysporum (No. 9), R. solani (No. 2) and S. rolfesii (No. 6) were found to be the most aggressive fungi on the base of the seedlings mortality. These pathogens were selected for further investigation.
Effect of elicitors on the linear growth of tested fungi: The tested elicitors significantly inhibited the linear growth of the isolated fungi. Rhizolex-T 50 and concentration of CHI (8 g L-1) prevented the growth of all tested fungi. The growth of F. oxysporum was standing in the treatments where SA and HQ at 1.4 and 1.2 g L-1 were found in the media, respectively (Table 2).
Greenhouse results: The efficacy of CHI, SA and HQ at the rate of 6 and 8, 0.7 and 1.4 and 0.6 and 1.2 g L-1, respectively, in form of seed soaking treatment and Rhizolex T-50 at concentration of 3 g kg-1, for controlling damping-off was evaluated under greenhouse condition.
Table 3 indicate that all tested elicitors significantly reduced pre-emergence damping-off syndromes compared with the check. However, SA at 0.7 g L-1 was insignificant in decreasing damping-off caused by F. oxysporum and R. solani. As for post-emergence, the treatments presented significant decreases than the check. CHI treatment at 8 g L-1 was superior in increasing healthy plants to reach (72.5, 80.9, 62.7 and 64.3% when F. solani, F. oxysporum, R. solani and S. rolfesii were found in the soil, respectively). The results show also that the fungicide (Rhizolex T-50) presented the highest reduction percentage in disease recovery.
| Table 1: | Frequency of the isolated fungi from roots of lupine plants showing root rot disease symptoms at different locations in Dakahlia Province |
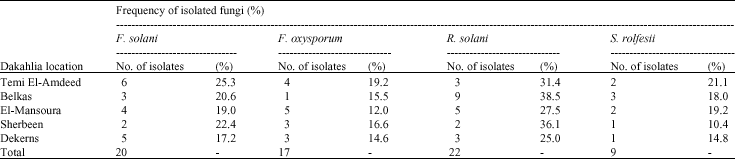 | |
| Samples were collected during 60 days of growing season | |
| Table 2: | Effect of the three elicitors on the linear growth (cm) of the most aggressive root rot fungi of lupine |
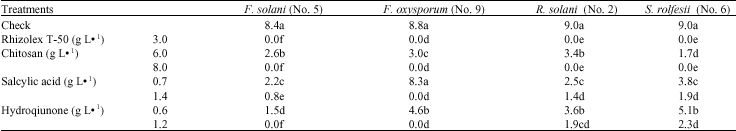 | |
| Values followed by the same letter(s) in each column do not differ significantly (p≤0.05) | |
| Table 3: | Effect of seed soaking in chitosan, salicylic acid, hydroquinone and Rhizolex T-50 on damping-off and seedling survivals of lupine, 40 days after sowing |
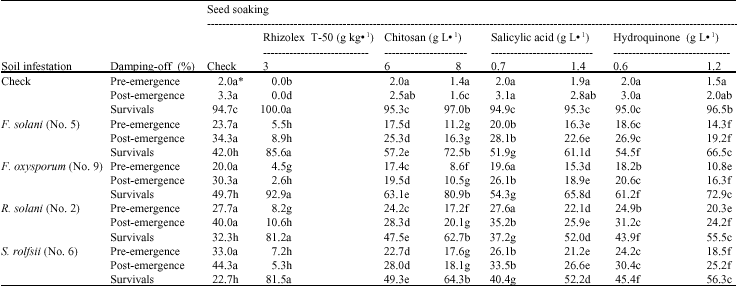 | |
| *Values followed by the same letter(s) in each row do not differ significantly (p≤0.05) | |
| Table 4: | Effect of the tested elicitors on lupine seedling survival from the root rot fungi under field conditions, 60 days after sowing |
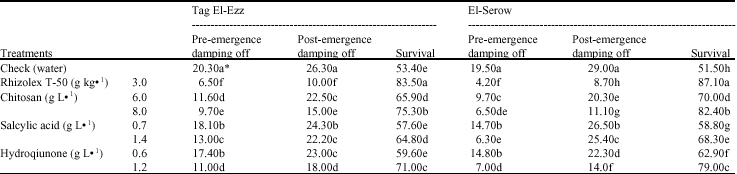 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05) | |
Field results
Disease assessment: Data presented in Table 4 show that soaking lupine seeds, in each one of the three elicitors or in the fungicide Rhizolex T-50, significantly decreased pre- and post emergence damping-off so, increased seedling survivals. CHI at 8 g L-1 came next to fungicide, in reducing lupine root rot disease. The percentage of reduction was 47.8 and 66.7%, while it was 87.98 and 78.46% in case of Rhizolex T-50 at pre-emergence stage and 43 and 61.7% and 61.98 and 70% at post-emergence damping-off stage, in Tag El-Ezz and El-Serow locations, respectively. HQ at 1.2 g L-1 came in the second order in reducing plant mortality and increasing lupine plant survivals as compared to check treatment. On the other hand, no significant difference was observed in plant survivals due to SA 0.7 or HQ 0.6 g L-1 applications as compared to check treatment.
Morphological characters: The effects of the three tested elicitors and the Rhizolex T-50 on plant height and number of branches and leaves plant-1 at 120 days from sowing are shown in Table 5. It was clear that the use of high levels of CHI (8 g L-1), HQ (1.2 g L-1) and SA (1.4 g L-1) gave the highest values of plant height. While, the low levels (6, 0.7 and 0.6 g L-1) as well as fungicide at concentration of 3 g kg-1 had no significant effect on the tested parameter. Concerning, number of branches and leaves, data reveal that Rhizolex T-50 as well as the tested elicitors significantly increased the tested parameters. The highest means occurred under the application of CHI at 8 g L-1 followed by HQ at 1.2 g L-1 then CHI at 6 g L-1. However, the fungicide came at the end compared with other treatments.
Yield and its components: Table 6 clearly show that the number of pods plant-1 was increased significantly when increasing elicitors concentrations. The highly significant increase occurred under the high concentration of CHI followed by HQ then SA. Application of CHI at 8 g L-1 caused a significant increase in the number of seeds and weight plant-1 as well as weight of 100-seeds. However, the other treatments had no significant effects in tested parameters except, the weight of 100-seeds, which showed significant increases when high concentration of HQ and SA were used.
Total phenol and photosynthetic pigments contents: Data in Table 7 reveal that Rhizolex T-50 and three tested elicitors increased the concentration of total phenols and the photosynthetic pigments in fresh leaves of lupine plants. The high concentration of total phenols occurred in the treatment where CHI 8 g L-1 was used, followed by fungicide with no significant differences. The recorded increases were 36.25 and 38.26% and 44.65 and 45.88%, respectively in Tag-zz and El-Serow localities. HQ at 1.2 g L-1 treatment cam in the second order in increasing lupine shoots content of total phenols as compared to check treatment.
| Table 5: | Effect of the tested elicitors on the growth parameters of lupine plant, 60 days after sowing |
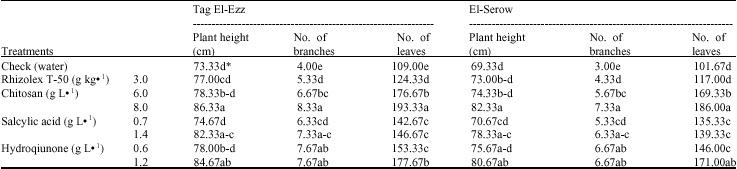 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05) | |
| Table 6: | Effect of elicitors on the yield components of lupine plant after 90 days from sowing |
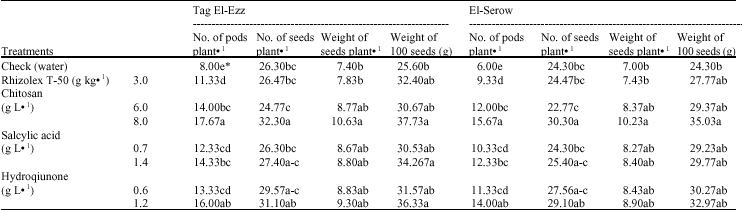 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05) | |
| Table 7: | Effect of the tested elicitors on phenol and photosynthetic pigment contents of lupine plant 60 days after sowing |
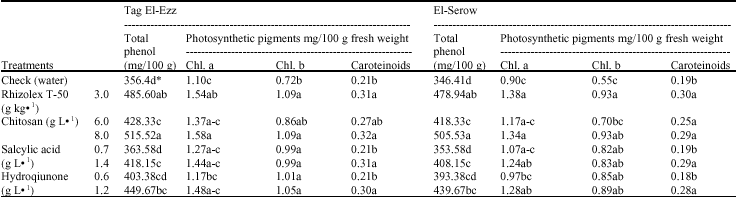 | |
| *Values followed by the same letter(s) in each column do not differ significantly (p≤0.05) | |
It was also observed that, chlorophyll a, b and caroteinoids concentration increased significantly in the application in which CHI 8 g L-1, fungicide and HQ 1.2 g L-1 and SA 1.4 g L-1 were applied. On the other hand, no significant differences were recorded between these treatments.
DISCUSSION
Identifying the defense activators (elicitors) that can supplement conventional chemical fungicides is a valuable contribution to lupine plants disease management, especially for the prevalence and destructive soil borne wilt and root rot pathogens. Isolation trials for the causal pathogens of lupine root rot symptoms showed that, R. solani and F. solani were the most dominant fungi isolated from infected roots in Dakahlia Province. Meanwhile, F. oxysporum and S. rolfesii were less frequent. From 68 fungal isolates, F. solani (No. 5), F. oxysporum (No. 9), R. solani (No. 2) and S. rolfsii (No. 6) were most aggressive in causing root rot of lupine seedlings. The effect of three elicitors (i.e., CHI, SA and HQ) on the linear growth of the isolated lupine root rot fungi was investigated in vitro. It was found an inverse relationship between the concentration of the elicitors and the linear growth of the tested fungi. A complete reduction of linear growth was observed when CHI at 8 g L-1 was tested against isolated fungi. HQ at 1.2 g L-1 and SA at 1.4 g L-1 were effective against F. oxysporum growth. These results are in harmony with the works demonstrated fungicidal activity of CHI against several fungi (Rabea et al., 2003; Hofgaard et al., 2005; Nawar, 2005; Hilal et al., 2006).
Nesci et al. (2003) introduced possible explanations on the antimicrobial action of elicitors including: (1) they may inhibit the functions of several enzymes by the oxidized compounds, possibly through reaction with sulfohydryl groups or through more nonspecific interactions with the proteins, (2) they dissolve in membrane lipids and interfere with membrane functions, including transport of nutrients, (3) they also interfere with the synthesis of protein, RNA and DNA, (4) they destroy the membrane potential similar to other week organic acids and/or (5) the site(s) and number of hydroxyl groups on the phenol compounds (salicylic acid and hydroquinone) are thought to be related to their relative toxicity to microorganisms, with evidence that increased hydroxylation associated with increased toxicity. In addition, (Scalbert, 1991) has found that the more highly oxidized phenols are the more inhibitory effect to the pathogen.
Soaking seeds of lupine in each tested elicitor had great values in decreasing (pre- and post-emergence) damping-off incidence, hence increasing survivals under greenhouse conditions. These positive results in decreasing lupine root rot severity were approved in two locations i.e., Dakahlia and Damietta Provinces. Several reports indicated that treating seeds of various crops with the elicitors such as CHI effectively controlled soil-borne diseases (Benhamou et al., 1998; Dasgupta et al., 1998; Jiang et al., 1999; Hilal et al., 2006). Field application of CHI for inducing resistance against late and early blight diseases of potato and root rot disease of lupine plants was reported by Abd-El-Kareem et al. (2001, 2004a). CHI was suggested to increase lignin biosynthesis and plant cell wall lignifications and effects on enzyme biosynthesis associated with the development of resistance (Tiuterev et al., 1996). Several studies have demonstrated that over expression of chitinases and B-1, 3-glucanase in plants is associated with enhanced resistance to various fungal pathogens (Chen et al., 1999; Abd-El-Karem et al., 2004b).
Soaked lupine seeds showed significant increase in lupine growth parameters, especially these soaked with CHI at 8 g L-1, followed by HQ at 1.2 g L-1. CHI was reported to enhance plant growth of other crops similar to the present results (Dasgupta et al., 1998; Hilal et al., 2006). These increases may be attributed to their effect on physiological processes in plant such as ion uptake, cell elongation, cell division, enzymatic activation and protein synthesis (Shakirova et al., 2003; Farouk, 2005; Amin et al., 2007).
Treatment of lupine seeds with CHI at 8 g L-1 and HQ at 1.2 g L-1 had led to increase in plant shoots and its content of total phenols. Phenolics are well-known as antifungal, antibacterial and antiviral compounds occur naturally in plants (Sivaprakasan and Vidhyasekaran, 1993). According to Matern and Kneusal (1988), the first step of the defence mechanism in plants involves a rapid accumulation of phenols at the infection site, which restricts or slows the growth of the pathogen. This effect might be due to the impact of these substances on enzymatic activity and translocation of the metabolites to lupine plant. The results presented here are in agreement with those obtained by Hilal et al. (2006) who reported that, total, free and conjugated phenols content in caraway and fennel leaves significantly increased by CHI application. This accumulation in phenolic compounds in leaves may be due to inhibition of catalase activity, which in turn induces phenylalanine lyase gene expression and synthesis of phenolic compounds (Vermerris and Nicholson, 2006). Yet total phenols have long been considered as important defense-related compounds whose levels are naturally high in resistant varieties of many crops (Gogoi et al., 2001; Khaleifa et al., 2006).
Chlorophyll content was determined in lupine plants since it is a good parameter reflects the health condition of plant. Noticeable increments in chlorophyll content were observed in plants developed from lupine seeds soaked in tested elicitors. This increment in chlorophyll content is by stimulating pigment formation and enhancing the efficacy of photosynthetic apparatus with a better potential for resistance and decrease in photophosphorylation rate usually occurring after infection (Amaresh and Bhatt, 1998). The earlier discussion may introduce an explanation for the high yield and growth parameters in lupine plants.
The increase in lupine yield may also be due to the role of elicitors in stimulation of physiological processes which reflect on improving vegetative growth that followed by active translocation of the photoassimilates. In this concern, SA might be regulating plant growth by increasing enzyme activity as α-amylase and nitrate reductase, which accelerate the sugar translocation from the leaves to developing fruit (Sharma et al., 1986). In addition, application of SA inhibits ethylene production leading to an increase in fruit number and consequently increases fruit yield per plant (Leslie and Romani, 1986).
This research recommended the use of different plant elicitors as alternative method for seed treatment and to avoid the hazards produced when using the toxic synthetic chemicals.
ACKNOWLEDGMENT
The authors thank Professor Mohamed Elwakil, Plant Pathology Department, Faculty of Agriculture, Mansoura University for critical review of manuscript.
REFERENCES
- Abd-El-Kareem, F., M.A. Abd-Alla and R.S.R. El-Mohamedy, 2001. Induced resistance in potato plants for controlling late blight disease under field conditions. Egypt. J. Phytopathol., 29: 29-41.
Direct Link - Abd-El-Kareem, F., M.A. Abd-Allah, N.G. El-Gamal and N.S. El-Mougy, 2004. Integrated control of Lupin root rot disease in solarized soil under greenhouse and field condition. Egypt. J. Phytopathol., 32: 49-63.
Direct Link - Abd-El-Karem, F., S. El-Mougy, Nehal, G. El-Gamal, Nadia and Y.O. Fatouh, 2004. Induction of resistance in squash plants against powdery mildew and alternaria leaf spot diseases using chemical inducers as protective or therapeutic treatments. Egypt. J. Phytopathol., 32: 65-76.
Direct Link - Chandra, A. and R.K. Bhatt, 1998. Biochemical and physiological response to salicylic acid in relation to the systemic acquired resistance. Photosynthetica, 35: 255-258.
CrossRef - Amin, A.A., E.S.M. Rashad and H.M.H. El-Abagy, 2007. Physiological effect of indole-3-butyric acid and salicylic acid on growth, yield and chemical constituents of onion plants. J. Applied Sci. Res., 3: 1554-1563.
Direct Link - Benhamou, N., P.J. Lafontaine and M. Nicole, 1994. Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan. Phytopathology, 84: 1432-1444.
CrossRefDirect Link - Benhamou, N., J.W. Kloepper and S. Tuzun, 1998. Induction of resistance against Fusarium wilt of tomato by combination of chitosan with an endophytic bacterial strain: Ultrastructure and cytochemistry of the host response. Planta, 204: 153-168.
CrossRefDirect Link - Chen, W.P., P.D. Chen, D.J. Liu, R. Kynast, B. Friebe and R. Velazhahan, 1999. Development of wheat scab symptoms is delayed in transgenic wheat plants that constitutively express a rice thaumatin-like protein gene. Theor. Applied Genet., 99: 755-760.
CrossRef - Eikemo, H., A. Stensvand and A.M. Tronsmo, 2003. Induced resistance as a possible means to control diseases of strawberry caused by Phytophthora spp. Plant Dis., 87: 345-350.
CrossRefDirect Link - Gogoi, R., D.V. Singh and K.D. Srivastava, 2001. Phenols as a biochemical basis of resistance in wheat against karnal bunt. Plant Pathol., 50: 470-476.
CrossRefDirect Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Hilal, A.A., M.G.A. Nada and W.H. Zaky, 2006. Induced resistance against Sclerotinia sclerotiorum disease in some Umbelliferous medicinal plants as a possible and effective control mean. Egypt. J. Phytopathol., 34: 85-101.
Direct Link - Hofgaard, I.S., A. Ergon, L.A. Wanner and A.M. Tronsmo, 2005. The effect of Chitosan and Bion on resistance to pink snow mould in perennial ryegrass and winter wheat. J. Phytopathol., 153: 108-119.
Direct Link - Khaleifa, M.M.A., C.R. Azzam and S.A. Azer, 2006. Biochemical markers associated with disease resistance to damping-off and root-rot diseases of peanut mutants and their productivity. Egypt. J. Phytopathol., 34: 53-74.
Direct Link - Leslie, C.A. and R.J. Romani, 1986. Salicylic acid: A new inhibitor of ethylene biosynthesis. Plant Cell Rep., 5: 144-146.
CrossRefDirect Link - Mackinney, G., 1941. Absorbtion of light by cholorophyll solution. J. Biol. Chem., 140: 315-322.
Direct Link - Matern, U. and R.E. Kneusel, 1988. Phenolic compounds in plant disease resistance. Phytoparasitica, 16: 153-170.
CrossRefDirect Link - Metraux, J.P., H. Signer, J. Ryals, E. Ward and M. Wyss-Benz et al., 1990. Increase in salicylic acid at the onset of systemic acquired resistance in cucumber. Science, 250: 1004-1006.
CrossRefPubMedDirect Link - Nawar, L.S., 2005. Chitosan and three Trichoderma spp. to control Fusarium crown and root rot of tomato in Jeddah, Kingdom of Saudi Arabia. Egypt. J. Phytopathol., 33: 45-58.
Direct Link - Nesci, A., M. Rodriguez and M. Etcheverry, 2003. Control of Aspergillus growth and aflatoxin production using antioxidants at different conditions of water activity and pH. J. Applied Microbiol., 95: 279-287.
CrossRefDirect Link - Oostendorp, M., W. Kunz, B. Dietrich and T. Staub, 2001. Induced disease resistance in plants by chemicals. Eur. J. Plant Pathol., 107: 19-28.
CrossRefDirect Link - Prapagdee, B., K. Kotchadat, A. Kumsopa and N. Visarathanonth, 2007. The role of chitosan in protection of soybean from sudden death syndrome caused by Fusarium solani f. sp. glycines. Bioresour. Technol., 98: 1353-1358.
CrossRefDirect Link - Rabea, E.I., M.E.T. Badawy, C.V. Stevens, G. Smagghe and W. Steurbaut, 2003. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules, 4: 1457-1465.
CrossRefDirect Link - Robinson, J.M. and S.J. Britz, 2000. Tolerance of a field grown soybean cultivar to elevated ozone level is concurrent with higher leaflet ascorbic acid level, higher ascorbate-dehydroascorbate redox status and long term photosynthetic productivity. Photosynth. Res., 64: 77-87.
CrossRefDirect Link - Scalbert, A., 1991. Antimicrobial properties of tannins. Phytochemistry, 30: 3875-3883.
CrossRefDirect Link - Shahda, W.T., 2000. The use of antioxidants for control of tomato damping-off. Alexandria J. Agric. Res., 45: 307-316.
Direct Link - Shakirova, F.M., A.R. Sakhabutdinova, M.V. Bezrukova, R.A. Fathudinova and D.R. Fathutdinova, 2003. Changes in the hormonal status of wheat seedlings induced by salicylic acid and salinity. Plant Sci., 164: 317-322.
CrossRefDirect Link - Sharma, S., S.S. Sharma and V.K. Rai, 1986. Reversal by phenolic compounds of abscisic acid-induced inhibition of in vitro activity of amylase from seeds of Triticum aestivum L. New Phytol., 103: 293-297.
CrossRefDirect Link - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link - Sticher, L., B. Mauch-Mani and J.P. Metraux, 1997. Systemic acquired resistance. Annu. Rev. Phytopathol., 35: 235-270.
CrossRefPubMedDirect Link