
S. Movassaghi
Laboratory of Cell Culture, Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran
G. Saki
Laboratory of Cell Culture, Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran
F. Javadnia
Laboratory of Cell Culture, Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran
M. Panahi
Laboratory of Cell Culture, Department of Anatomy, School of Medicine, Jondishapour University of Medical Sciences, Ahwaz, Iran
M. Mahmoudi
Department of Biostatistics, School of Medicine, Tehran University of Medical Sciences, Tehran, Iran
F. Rhim
Physiology Research Center, Jondishapour University of Medical Sciences, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 1 | Page No.: 19-25







ABSTRACT
MBCD and Cholesterol-Loaded-Cyclodextrin (CLC) were examined for their abilities to increase the cryosurvival of C57BL/6 mouse sperm, the main strain of genetically engineered mice. The intactness of acrosome and motility of frozen/thawed spermatozoa were used to monitor cryosurvival. In this experimental study, male mice were randomly divided in 6 groups: control 1, experimental 1, experimental 2, control 2, experimental 3 and experimental 4. In experimental groups 1 and 2 spermatozoa were exposed to 0.75 and 1 mM MBCD and in experimental groups 3 and 4 were exposed to two different concentrations of CLC (1 and 2 mg mL-1) over a period of 1 h and were subsequently cryopreserved. Spermatozoa in control 1 group were frozen without any exposure to CLC or MBCD and in control 2 (vehicle), sperms were incubated with 4 mM MBCD. The post-thaw sperms were evaluated for their motility and acrosomal status. The values of the intact acrosome and motility increased significantly with concentration of CLC compared to controls and MBCD experimental groups (p<0.05). These results indicate that cryosurvival of C57BL/6 mouse spermatozoa is enhanced by exposure to MBCD which loaded with cholesterol (CLC) before freezing and MBCD alone can not protect sperm from freeze-thaw damage efficiently compare to CLC.
PDF Abstract XML References Citation
How to cite this article
S. Movassaghi, G. Saki, F. Javadnia, M. Panahi, M. Mahmoudi and F. Rhim, 2009. Effects of Methyl-Beta-Cyclodextrin and Cholesterol on Cryosurvival of Spermatozoa from C57BL/6 Mouse. Pakistan Journal of Biological Sciences, 12: 19-25.
DOI: 10.3923/pjbs.2009.19.25
URL: https://scialert.net/abstract/?doi=pjbs.2009.19.25
DOI: 10.3923/pjbs.2009.19.25
URL: https://scialert.net/abstract/?doi=pjbs.2009.19.25
INTRODUCTION
Since the routine introduction of frozen semen in the dairy industry 50 years ago, the cryopreservation of gametes has been widely developed. Cryopreservation of spermatozoa has been great benefit to agriculture, aquaculture, biotechnology and the conservation of wild animals and the treatment of infertility in human reproduction (Zeng and Terada, 2001) and provides a much simpler and more economical alternative to the freezing of embryos for the storage of genetically engineered strains of mice in facilities and research laboratories. In general, relatively high fertilization rates are obtained for frozen/thawed sperm of CBA/JN, DBA/2N and C3H inbred strains and some F1 hybrid strains. However, rates are remarkably low in frozen/thawed sperm of C57BL/6 mice, the main strain used not only for the production of transgenic mice but also as a backcross for the targeted mutant mice (Takeo et al., 2008).
However, if the freezing and thawing process occurs without any protective treatment of the sperm membrane, it usually results in significant damage to the membrane (Zeng and Terada, 2001; Chakrabarty et al., 2006; Waston, 1995; Watson, 2000). Even when successful cryopreservation protocol is used, about half of the spermatozoa are killed or immobilized during freezing and thawing. Then it is necessary to understand sperm preservation mechanisms especially at the molecular level, to determine how sperm membranes respond to freezing and thawing (Zeng and Terada, 2001). Altering the lipid composition of plasma membranes not only affects the ability of sperm to capacitate and acrosome react, but also affects the way sperm respond to cryopreservation. When cyclodextrins, cyclic oligosaccharides of glucose that contain a hydrophobic center capable of incorporating lipids, are preloaded with cholesterol to form Cholesterol-Loaded-Cyclodextrin (CLC) and then incubated with bull sperm before cryopreservation, higher percentages of motile and viable cells are recovered after freezing and thawing, compared with control sperm (Purdy and Graham, 2004; Combes et al., 1998). This added cholesterol most likely benefits cells by eliminating or at least lowering the temperature at which the sperm plasma membranes undergo the lipid phase transition from the fluid to the gel state as the cells are cooled.
Several observations suggest that CLC is capable of improving cryosurvival of frozen/thawed sperm of some mammalian species (Purdy and Graham, 2004; Combes et al., 1998; He et al., 2001; Moore et al., 2005; Purdy et al., 2005; Galantino-Homer, 2006; Torres et al., 2006; Moce and Graham, 2006; Li et al., 2006). The amount of cholesterol is important in maintaining membrane integrity during cryopreservation and CLC with altering the lipid composition of sperm plasma membranes can affect the cryosurvival of this cell. However, there has been no report about the effect of CLC on cryosurvival of mouse sperm.
On the contrary, some researches indicate that membrane cholesterol efflux induces an enhanced membrane fluidity and permeability (Grunze and Deuticku, 1974; Cooper et al., 1978) and the elevation of membrane cholesterol is associated with decreased membrane fluidity (Cooper et al., 1978; Vanderkooi et al., 1974; Shattil and Cooper, 1976; Madden and Quinn, 1979; Hartel et al., 1998). Zeng and Terada (2001) found that cryosurvival of boar spermatozoa is enhanced by exposure to MBCD before freezing. The effect of MBCD and cholesterol (CLC) on cryosurvival of spermatozoa was examined in two different phases of experiments: 1st phase Experiment and 2nd phase experiment.
MATERIALS AND METHODS
Animals: This experimental study was performed in Cell Culture Laboratory of Ahwaz Jondishapour University of Medical Sciences (AJUMS). C57BL/6 male mice (with 12-week-old age) were purchased from AJUMS animal research center and used as donors of sperm. The fertilizing ability of male mice was proved before to starting the experiments. They were housed singly or at maximum of two per cage for at least 5 days before sperm collection. Animals were kept under a 12 h/12 h dark/light cycle at a 22±1°C temperature with free access to food and water All mice were kept according to the guidelines for the care and use of laboratory animals.
Materials: All chemicals were reagent grade and purchased from Sigma (St. Louis, MO), except for antifade solution, which purchased from Molecular Probes (Eugene, OR).
Cryoprotectant solution and culture media: The cryoprotectant solution (CPA), containing 18% raffinose pentahydrate and 3% skim milk in distilled water was prepared according to the method described by Nakagata and Takeshima (1993). After warming (60°C) for total dissolution of the sugar, the CPA was centrifuged at 10,000 x g for 10 min. The supernatant was filtered through a 0.45 μm filter and solution was stored at -20°C. T6 solution without bovine serum albumin or BSA (in order to clearly test the effect of MBCD) was used as a medium for sperm preincubation. Three various concentrations of MBCD (0.75, 1 and 4 mM) were dissolved in T6 medium at room temperature.
CLC preparation: As earlier described by Zeng and Terada (2001) cholesterol-3-sulfate with two concentrations (1 and 2 mg mL-1) was added to T6 media containing 4 mM MBCD, then sonicated using Ultrasonic Disrupture and filtered through a 0.22 μm filter. This solution was prepared freshly.
Sperm collection: Male mice in control 1 group were killed by cervical dislocation. Both cauda epididymis were removed and each of them was cut seven times with the edge of a 30-gauge needle and placed in a sterile plastic vial (Eppendorf) containing 1 mL of CPA equilibrated beforehand at 37°C. Sperm were allowed to disperse (swim out) for 10 min in the CPA solution, then the tissue was removed. Each sample was obtained from one cauda epididymis. In experimental groups 1 and 2 and control 2 group the tissues were put in T6 media containing 0.75, 1 and 4 mM MBCD, respectively. In CLC treated groups, cauda epididymis was submerged in T6 media containing two different concentrations of CLC (1 mg mL-1 CLC; experiment group 1 and 1 mg mL-1 CLC; experimental group 2) and after swim out, spermatozoa were incubated for 60 min under 5% CO2 in air at 37°C. Then the sample was centrifuged at 735 x g for 4 min. The supernatant was discarded and replaced with 1 mL CPA.
Sperm analysis: Concentration, progressive motility and total motility of the frozen/thawed sperm samples were determined using a Makler chamber. Every sperm sample was analyzed twice. All counts were performed at 37°C in T6 media. Total motility was defined as any movement of the sperm head and progressive motility was defined as the count of those spermatozoa that moved in a forward direction.
Freezing and thawing procedure: Sperm samples were distributed in aliquots of 100 μL into nine 1.8 mL cryotubes (Nunc Cryotubes, Denmark). After capping, vials were immediately placed in the vapor phase of a liquid nitrogen storage container for 10 min. After that time, the tubes were plunged into the liquid nitrogen (-196°C) for storage. After 5 days, frozen samples were thawed by transferring them from liquid nitrogen into a 37°C water bath for 2 min (Sztein et al., 2000). The thawed sperm suspension was incubated for 30 min with 5% CO2 in air at 37°C in a 200 μL drop of T6 medium (Nishizono et al., 2004).
Assessing acrosomal status by FITC-PNA staining: For acrosome staining, a procedure described before was used (Zeng and Terada, 2001; Fazeli et al., 1997). Briefly, 30 μL of frozen/thawed sperm samples were smeared onto microscope slides, air-dried and fixed with absolute methanol for 10 min in -20°C. Thirty microliter of fluorescein isothiocyanate- peanut agglutinin (FITC-PNA) solution (100 μg mL-1) in PBS were spread over each slide. The slides were then incubated in a dark, moist chamber for 30 min at 37°C. They were subsequently rinsed with PBS and air-dried, then mounted with 10 μL of antifade solution to preserve fluorescence. A cover slip was then applied and the edges were sealed with colorless nail polish.
The acrosome status of spermatozoa was monitored and photographed with an epifluorescence microscope (BH2; Olympus, Japan). One hundred cells per slide were counted. All samples were coded before evaluation and were evaluated by one observer. The fluorescence images could be classified into 3 groups: (1) spermatozoa with intensively bright fluorescence of the acrosomal cap, indicating an intact acrosome; (2) spermatozoa with disrupted fluorescence of the acrosomal cap, indicating partially damaged acrosome and (3) spermatozoa with no fluorescence, indicating a damaged acrosome. The last group was identified under bright field illumination of the microscope.
Experimental design: The animals were put in six groups randomly: control 1 (without MBCD or CLC), experimental group 1 (MBCD = 0.75 mM), experimental group 2 (MBCD = l mM), control 2 (MBCD = 4 mM), experimental group 3 (CLC = 1 mg mL-1) and experimental group 4 (CLC = 2 mg mL-1). The effect of MBCD on cryosurvival of spermatozoa was examined with two experiments.
Ist phase: Spermatozoa were incubated in T6 media supplemented with various concentrations of MBCD alone or the combination of MBCD and cholesterol (CLC) for 60 min and then frozen. After 5 days the samples were thawed and the motility of frozen/thawed sperm determined.
2nd phase: After thawing, the acrosomal status of samples was studied with FITC- PNA staining and fluorescence microscope.
Statistical analysis: All percentage data were subjected to arcsine transformation. The statistical analysis was performed using SPSS version 13.0.1 software (SPSS Inc, USA). Data were analyzed by ANOVA and statistical differences between the various treatment group means were determined by Tukey test. Data are given as the Mean±SD. Differences between the means were considered to be significant when p<0.05 was achieved.
RESULTS
Ist phase
Effect of preincubation with MBCD and CLC on the motility of cryopreserved mouse spermatozoa: Motility of spermatozoa in control 2 group was significantly lower than other groups before freezing and after thawing (Table 1). But there were no significant differences in motility of frozen/thawed spermatozoa between control 1 and experimental groups 1 and 2 (p<0.05). The percentage of total motile sperm after thawing was increased greatly when the concentration of CLC was increased. There were significant differences (p<0.05) in total motility between control 1 group (29.14%) and experimental groups 3 and 4 that preincubated with CLC (65.95 and 71.45%, respectively) before freezing. Overall, exposure to CLC before freezing strongly increased the motility of frozen/thawed spermatozoa, this effect is increased when the concentration of CLC is increased and the better effect was achieved with 2 mg mL-1 CLC.
2nd phase
Effect of preincubation with MBCD and CLC on the acrosomal status of cryopreserved mouse spermatozoa: The percentage of post-thaw spermatozoa with intact acrosomes was greatly increased (p<0.05) by the addition of CLC. When treated with 2 mg mL-1 CLC, the highest percentage of post-thaw spermatozoa possessing intact acrosomes was noted (43.73%). In contrast, the percentage of spermatozoa with damaged acrosomes was found to reduce under CLC treatments (group 1 = 15.14%, group 2 = 13.5%) as compared with the control group (57.32%). But when T6 media supplemented with 0.75 mM, 1 mM or 4 mM MBCD (control 2), led to decrease (p<0.05) in percentage of post-thaw spermatozoa with intact acrosomes (Table 2). These results indicate that sperm acrosome gains strong protection against cryoinjury by incubation of sperm with CLC before freezing process.
| Table 1: | Effect of MBCD and MBCD+cholesterol on total motility of spermatozoa before freezing and after thawing in control and experimental groups |
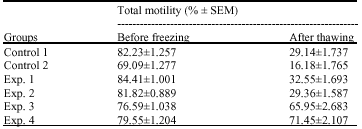 | |
| Table 2: | Effect of MBCD and MBCD+cholesterol on acrosome status of spermatozoa after thawing in control and experimental groups |
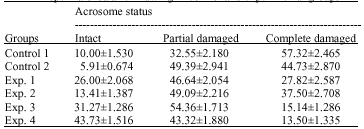 | |
DISCUSSION
Cryopreservation exposes sperm to mechanical and anisosmotic stresses that reducing cell survival and alter surviving sperm function, in turn, reducing cell longevity and fertility compared with fresh sperm (Moce and Graham, 2006).
Sperm sensitivity to cold shock damage is determined by the membrane phospholipids composition and the membrane cholesterol to phospholipids ratio (Holt, 1997). Sperm possessing high cholesterol to phospholipids ratios are more resistant to cold shock damage than sperm having low cholesterol to phospholipids ratios (Moce and Graham, 2006).
Cyclodextrins can be used to alter the cholesterol content of cell membranes (Christian et al., 1997; Visconti et al., 1997). They are able to mediate sperm membrane cholesterol efflux efficiently (Zeng and Terada, 2001; Visconti et al., 1999) and if cyclodextrins are preloaded with cholesterol they insert cholesterol into membranes (Navratil et al., 2003). As mentioned before, many observations suggest that depletion of cholesterol from cell membrane increases membrane fluidity and permeability (Zeng and Terada, 2001; Grunze and Deuticke, 1974; Cooper et al., 1978). If the cell is sufficiently permeable to water during freezing, the differential vapor pressure for water across the plasma membrane would remain small and rapid dehydration would be induced as water moves out of the cell in accordance with extracellular freezing. During cryopreservation, avoidance of numerous and large intracellular ice crystals is necessary for cell survival (Gao et al., 1997). In most cases, cells are killed due to undergoing intracellular ice formation. Increasing the membrane fluidity and permeability may reduce the amount of intracellular ice formation and therefore minimize the freeze-thaw damage (Zeng and Terada, 2001). Some researches indicate that cyclodextrins with decreasing the amount of sperm membrane cholesterol, increase the membrane fluidity and improve cryosurvival of boar spermatozoa in terms of intact acrosome and motion parameters and have protective effects on sperm against cold shock (Zeng and Terada, 2001).
On the contrary, many researches showed that if stallion, bull or ram sperm are treated with CLC before freezing, they exhibit greater cryosurvival rates than untreated sperm (Purdy and Graham, 2004; Combes et al., 1998; He et al., 2001; Moore et al., 2005; Purdy et al., 2005; Galantino-Homer, 2006; Torres et al., 2006; Moce et al., 2006; Li et al., 2006). The exact mechanism by which added cholesterol protects sperm membranes is not known. As stated earlier, species with high cholesterol to phospholipids ratios are resistant to cold shock. At least part of the sperm damage induced from cold shock is due to lipid phase transition that the membrane experiences during the cooling process. High cholesterol levels stabilize membranes during cooling. The cholesterol content in the membranes of bull and stallion sperm increased 2 to 3 fold compared with control sperm after treatment with CLC (Purdy and Graham, 2004; Moore et al., 2005) and it remains greater than in control sperm after cryopreservation (Moore et al., 2005). This increased cholesterol content in bull and stallion sperm raised the cholesterol to phospholipids ratio in these to values similar to those sperm that are cold-shock resistant (Purdy and Graham, 2004; Moore et al., 2005). It is likely that the lipid phase transition is eliminated or the temperature at which it occurs is lower for CLC treated bull and stallion sperm than for control sperm. Supporting this idea, Purdy et al. (2005) demonstrated increased membrane fluidity at lower temperatures for bull sperm treated with CLC than for untreated sperm. On the other hand, cholesterol could be increasing sperm membrane permeability to cryoprotectants and lessening osmotic cell damage, because CLC treatment increases the osmotic tolerance of stallion sperm (Moore et al., 2005). This is very important because sperm experience large volume changes when cryoprotectants are added or removed and their membranes can suffer damage during these process (Moce and Graham, 2006). In the present study, we have shown that CLC improves the cryosurvival of frozen/thawed C57BL/6 mouse sperm, too. In contrast, MBCD alone has no beneficial effect on cryosurvival of mouse spermatozoa.
Treating mouse sperm with CLC resulted in increased percentage of total motile sperm (group 1 = 65.95%, group 2 = 71.45%) compared with control group (29.14%) that increased with concentration of CLC. Moore et al. (2005) reported that differences in motility between CLC-treated and untreated sperm are due to changes in osmotic tolerance and CLC treatment increases the osmotic tolerance limits of stallion sperm compared with the control samples (Moore et al., 2005). Li et al. (2006) and Purdy and Graham (2004) then also reported an increase the percentage of motile sperm after thawing for bull sperm that had been treated with CLC before cryopreservation.
Findings of Nishizono et al. (2004) showed that the acrosome contents were missing from frozen sperm of C57BL/6 mouse spermatozoa. The acrosome contents are vital proteins for passing through the zona pellucida surrounding an oocyte at fertilization, especially acrosin but it lost during cryotreatment. Muller et al. (1999) also reported that the plasma membrane of the acrosome was changed and the acrosome contents were reduced in frozen ram spermatozoa. These results suggest that frozen/thawed C57BL/6 mouse spermatozoa can not induce an acrosomal reaction and can not penetrate the zona pellucida of the egg. Thus, the fertilization ability of mouse sperm is lost during cryotreatment (Nishizono et al., 2004). One of the initial steps in sperm capacitation is a loss of cholesterol from the plasma membrane (Ehrenwald et al., 1988; Langalis and Roberts, 1985). This cholesterol efflux induces plasma membrane lipid reorganization, ultimately increasing membrane permeability to Ca2+, HCO3- and K+ (Visconti and Kopf, 1998). High intracellular concentrations of these ions are required for a spermatozoon to undergo the acrosomal reaction as well as fuse with oocyte. The time at which a particular sperm initiates capacitation and acrosomal reaction depend in large part on a cell`s membrane status and in particular on the amount of cholesterol contained in the plasma membrane. During capacitation, when sufficient cholesterol is removed, the plasma membrane becomes unstable, enhancing its ability to fuse with the outer acrosomal membrane, resulting in the acrosomal reaction. Sperm capacitation can be retarded by adding lipid vesicles composed of synthetic phospholipids liposome containing cholesterol to the sperm as these prevent the loss of cholesterol from the membrane. Purdy and Graham (2004) reported that increasing the cholesterol content of the plasma membrane would retard sperm capacitation and acrosomal reaction. On the other hand, this added cholesterol prevents sperm plasma membrane to undergo the early acrosome reaction and loss of acrosomal contents during cryopreservation. It is generally accepted that the slighter the acrosomal damage during cryopreservation, the higher level of fertility that will be established (Zeng and Terada, 2001). In the present study, the percentage of spermatozoa with intact acrosomes increased greatly with the aid of CLC treatment and the highest was obtained in the presence of 2 mg mL-1 CLC. Exposure to this concentration of CLC also yielded a significant higher motility compared with controls. In agreement with these results. Moore et al. (2005) reported that addition of CLC increased the percentage of acrosome intact stallion sperm after cryopreservation compared to untreated sperm. Galantino-Homer et al. (2006) also reported the same effect of CLC on porcine sperm. But the results of another studies showed that CLC decreased the percentage of boar spermatozoa with intact acrosome as well as sperm motility (Zeng and Terada, 2001). The results conflict with those in the present study. This discrepancy may be due to differences in the experimental conditions and the species that have been used (boar and mouse).
In conclusion, this study suggests that exposure to CLC is strongly beneficial to cryosurvival of C57BL/6 mouse sperm and provides new information to modify the cryopreservation method of mouse sperm. However, this is the first report about the effect of MBCD on cryosurvival of C57BL/6 mouse sperm and further investigations are necessary.
ACKNOWLEDGMENT
This study was supported by the Vice-Chancellor for research of Ahwaz Jondishapour University of Medical Sciences.
REFERENCES
- Chakrabarty, J., D. Banerjee, D. Pal, J. De and A. Ghosh et al., 2007. Shedding off specific lipid constituents from sperm cell membrane during cryopreservation. Cryobiology, 54: 27-35.
CrossRefPubMedDirect Link - Cooper, R.A., M.H. Leslie, S. Fischkoff, M. Shinitzky and S.J. Shattil, 1978. Factors influencing the lipid composition and fluidity of red cell membranes in vitro: Production of red cells possessing more than two cholesterol per phospholipids. Biochemistry, 17: 327-331.
PubMedDirect Link - Christian, A.E., M.P. Haynes, M.C. Phillips and G.H. Rothblat, 1997. Use of cyclodextrins for manipulating cellular cholesterol content. J. Lipid Res., 38: 2264-2272.
PubMed - Ehrenwald, E., R.H. Foote and J.E. Parks, 1988. Cholesterol efflux from bovine sperm.І. Induction of the acrosome reaction with lysophosphatidylcholine after reducing sperm cholesterol. Gamete Res., 20: 145-157.
CrossRefPubMedDirect Link - Fazeli, A., W.J. Hage, F.P. Cheng, W.F. Voorhout and A. Marks et al., 1997. Acrosome intact boar spermatozoa initiate binding to the homologous zona pellucida in vitro. Biol. Reprod., 56: 430-438.
PubMed - Galantino-Homer, H.L., W.X. Zeng, S.O. Megee, M. Dallmeyer and D. Voelkl et al., 2006. Effect of 2-hydroxypropyl-beta-cyclodextrin and cholesterol on porcine sperm viability and capacitation status following cold shock or incubation. Mol. Reprod. Dev., 73: 638-650.
CrossRefDirect Link - Grunze, M. and B. Deuticke, 1974. Changes of membrane permeability due to extensive cholesterol depletion in mammalian erythrocytes. Biochim. Biophys. Acta., 356: 125-130.
PubMed - Hartel, S., H.A. Diehl and S.F. Ojeda, 1998. Methyl-beta-cyclodextrins and liposomes as water-soluble carriers for cholesterol incorporation into membranes and its evaluation by microenzymatic fluorescence assay and membrane fluidity-sensitive dyes. Anal. Biochem., 258: 277-284.
PubMed - He, L., J.L. Bailey and M.M. Buhr, 2001. Incorporating lipids into boar sperm decreases chilling sensitivity but not capacitation potential. Biol. Reprod., 64: 69-79.
PubMedDirect Link - Holt, W.V., 1997. Alternative strategies for long-term preservation of spermatozoa. Reprod. Fertil. Dev., 9: 309-319.
PubMed - Langalis, J. and K.D. Roberts, 1985. A molecular membrane model of sperm capacitation and the acrosome reaction of mammalian spermatozoa. Gamete Res., 12: 183-224.
CrossRef - Li, G., J. Saenz, R.A. Godke and R.V. Devireddy, 2006. Effect of glycerol and cholesterol-loaded cyclodextrin on freezing-induced water loss in bovine spermatozoa. Reproduction, 131: 875-886.
CrossRefDirect Link - Moce, E. and J.K. Graham, 2006. Cholesterol-loaded cyclodextrins added to fresh bull ejaculates improve sperm cryosurvival. J. Anim. Sci., 84: 826-833.
PubMed - Moore, A.I., E.L. Squires and J.K. Graham, 2005. Adding cholesterol to stallion sperm plasma membrane improves cryosurvival. Cryobiology, 51: 241-249.
CrossRefDirect Link - Muller, K., T. Pomorski, P. Muller and A. Hermann, 1999. Stability of transbilayer phospholipid asymmetry in viable ram sperm cells after cryotreatment. J. Cell Sci., 112: 11-20.
PubMed - Muller, K., P. Muller, G. Pincemy, A. Kurz and C. Labbe, 2008. Characterization of sperm plasma membrane properties after cholesterol modification: Consequence for cryopereservation of rainbow trout spermatozoa. Biol. Reprod., 78: 390-399.
PubMed - Nakagata, N. and T. Takeshima, 1993. Cryopreservation of mouse spermatozoa from inbred and F1 hybrid strains. Jikken. Dobutsu., 42: 317-320.
PubMed - Navratil, A.M., S.P. Bliss, K.A. Berghorn, J.M. Haughian and T.A. Farmerie et al., 2003. Constitutive localization of the gonadotropin-releasing hormone (GnRH) receptor to low density membrane microdomains is necessary for GnRH signaling to ERK. J. Biol. Chem., 278: 31593-31602.
CrossRefPubMedDirect Link - Nishizono, H., M. Shioda, T. Takeo, T. Irie and N. Nakagata, 2004. Decrease of fertilizing ability of mouse spermatozoa after freezing and thawing is related to cellular injury. Biol. Reprod., 71: 973-978.
CrossRefPubMedDirect Link - Purdy, P.H., M.H. Fox and J.K. Graham, 2005. The fluidity of Chinese hamster ovary cell and bull sperm membranes after cholesterol addition. Cryobiology, 51: 102-112.
CrossRefPubMedDirect Link - Purdy, P.H. and J.K. Graham, 2004. Effect of adding cholesterol to bull sperm membranes on sperm capacitation, the acrosome reaction and fertility. Biol. Reprod., 71: 522-527.
CrossRefDirect Link - Sztein, J., J. Farley and L. Mobraaten, 2000. In vitro fertilization with cryopreserved inbred mouse sperm. Biol. Reprod., 63: 1774-1780.
PubMedDirect Link - Shattil, S.J. and R.A. Cooper, 1976. Membrane microviscosity and human platelet function. Biochemistry, 15: 4832-4837.
PubMed - Takeo, T., T. Hoshii, Y. Kondo, H. Toyodome and H. Arima et al., 2008. Methyl-beta-cyclodextrin improves fertilizing ability of C57BL/6 mouse sperm after freezing and thawing by facilitating cholesterol efflux from the cells. Biol. Reprod., 78: 546-551.
CrossRefPubMedDirect Link - Torres, P., C. Serres, C. Gomez-Cuetara, I. Santiago and E. Mateos et al., 2006. Effect of cholesterol loaded cyclodextrin on motility and plasma membrane integrity of cooled stallion sperm. Anim. Reprod. Sci., 94: 148-151.
CrossRef - Vanderkooi, J., S. Fischkoff, B. Chance and R.A. Cooper, 1974. Fluorescent probe analysis of architecture of natural and experimental cholesterol-rich membrane. Biochemistry, 13: 1589-1595.
PubMed - Visconti, P.E. and G.S. Kopf, 1998. Regulation of protein phosphorylation during sperm capacitation. Biol. Reprod., 59: 1-6.
PubMed - Visconti, P.E., H. Galantino- Hommer, X. Ning, G.D. Moore and J.P. Valenzuela et al., 1999. Cholesterol efflux-mediated signal transduction in mammalian sperm. J. Biol. Chem., 274: 3235-3242.
PubMed - Watson, P.F., 1995. Recent developments and concepts in the cryopreservation of spermatozoa and the assessment of their post-thawing function. Reprod. Fertil. Dev., 7: 871-891.
CrossRefPubMedDirect Link - Woods, E.J., J.D. Benson, Y. Agca and J.K. Critser, 2004. Fundamental cryobiology of reproductive cells and tissues. Cryobiology, 48: 146-156.
CrossRefPubMedDirect Link - Zeng, W.X. and T. Terada, 2001. Effect of methyl-beta-cyclodextrin on cryosurvival of boar spermatozoa. J. Androl., 22: 111-118.
PubMedDirect Link - Zeng, W.X. and T. Terada, 2001. Protection of boar spermatozoa from cold shock damage by 2-Hydroxypropyl-beta-cyclodextrin. Theriogenology, 55: 615-627.
PubMedDirect Link - Combes, G.B., D.D. Varner, F. Schroeder, R.C. Burghardt and T.L. Blanchard, 1998. Effect of cholesterol on the motility and plasma membrane integrity of frozen equine spermatozoa after thawing. Proceedings of the 7th International Symposium on Equine Reproduction, July 12-17, 1998, Pretoria, South Africa, pp: 35-36.
- Nishizono, H., M. Shioda, T. Takeo, T. Irie and N. Nakagata, 2004. Decrease of fertilizing ability of mouse spermatozoa after freezing and thawing is related to cellular injury. Biol. Reprod., 71: 973-978.
CrossRefPubMedDirect Link