
M. Safari
Department of Anatomy, Semnan University of Medical Science, Semnan, Iran
L. Ghahari
Department of Anatomy, Artesh University of Medical Science, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 16 | Page No.: 1156-1159







ABSTRACT
The aim of this study was to study the changes of inducible and constitutive Nitric Oxide Synthase (NOS) after brain injury. In order to brain injury 42 wistar rats were submitted and divided in 7 groups. Nitric oxide synthase activities were assayed at different times after injury. Present results showed that a significant increase of iNOS and cNOS activity 8 h after lesion. In conclusion, both isoformes of NOS increase at different time after brain injury.
PDF Abstract XML References Citation
How to cite this article
M. Safari and L. Ghahari, 2009. Alterations of Inducible and Constitutive Nitric Oxide Synthase after Hippocampal Injury in Rats. Pakistan Journal of Biological Sciences, 12: 1156-1159.
DOI: 10.3923/pjbs.2009.1156.1159
URL: https://scialert.net/abstract/?doi=pjbs.2009.1156.1159
DOI: 10.3923/pjbs.2009.1156.1159
URL: https://scialert.net/abstract/?doi=pjbs.2009.1156.1159
INTRODUCTION
Traumatic Brain Injury (TBI) induces a complex cascade of pathophysiological changes that spread from primary lesion to the adjacent tissues (Ziaja et al., 2007). Some process in CNS damage includes, ischemia, inflammatory responses, alteration of energic metabolism. As a result some important amino acids such as glutamate, aspirate are altered after damage and this lead to disregulation of some metabolic pathway (Diaz-Ruiz et al., 2007). Among the important free radicals that produced during brain damage is Nitric Oxide (NO). NO is synthesized from L-arginin by two classes of enzymes termed constitutive nitric oxide Synthase (cNOS) and inducible (iNOS), the former dependent on intra cellular calcium and the latter independent. In the CNS two different cNOS isoformes are present, one in neurons (nNOS) and in endothelial cells (eNOS) (Hierholzer et al., 1998; Martinez-Murillo et al., 2007). CNS and neural cells have a great Ca2+/calmodolin regulated NOS activity and NO is an important biological messenger in the CNS (Matsumoto et al., 1993). NOS have been purified from microglia (Dinerman et al., 1994), cerebellum, hippocampus (Gham et al., 2002), striatum (Sardo et al., 2002). It has been suggested that NO may play a role in learning, memory and development of the brain (Delwing et al., 2008). There is a little information about NOS activity after brain injury (Diaz-Ruiz et al., 2002). Diazal-Ruiz et al. (2002) showed a significant increase in cNOS activity at 4 and 8 h and iNOS activity was increased 72 h after spinal cord injury (Hu et al., 2008). Hu et al. (2008) showed after spinal cord injury a time-dependent increase in cNOS and nNOS activity. Xu et al. (2000) showed after traumatic injury of CNS the activity of iNOS increased. Cherian et al. (2000) reported that after brain injury in rats pretreated with L arginin concentration of NO were increased. We hypothesized that enhanced NO production resulting from iNOS and eNOS expression would contribute to inflammatory signaling in brain damage. In the present study, we inflicted a contusion to the brain cause a changes of cNOS and iNOS activities in rats.
MATERIALS AND METHODS
Animal: The animal experiments were conducted in Pharmacology and Anatomy Department and NOS measurement works were conducted in cellular and molecular research center of Iran medical university from 2006-2008.
Adult male Wistar strain of rats (about 3 months of age) were used and were housed in a cages with a 12 h light/dark cycle at 22-24°C temperature, with food and water adlibitum. All experimental procedures were carried out in accordance with the National Institutes of Health guide for the care and use of laboratory animals and all experiments conformed to Iran University of medical sciences guidelines on the ethical use of laboratory animals. Also, in each experiment care was taken to minimize the animals suffering and to use the minimum number of animals.
Hippocampus injury: The rats were randomly allocated in two groups. One group (lesion group n = 6 in each subgroup) was submitted to a sever hippocampal injury according with the injection of a canola in to the right hippocampus with the stereotaxic apparatus (3.8 mm from bregma, 2.6 mm from the mid line and 3.7 mm vertically) according to atlas of the rat brain (Paxinos and Watson, 2005).This coordination has been used effectively in several studies for the hippocampus. The second group was only submitted to the drain of skull (sham group n = 6) under pentobarbital anesthesia.
Measurement of NO: NOS activity were assayed by measuring the conversion of [3H] - L-arginin to the [3H] - L-Citrulline (Bredt and Synder, 1990). Both groups allowed to recover from anesthesia and surgical procedures, then housed in individual cages. Rats take food and water ad libitum.
Animal were killed at different time after surgery (2, 4, 6, 8, 12, 24 and 72 h). After decapitation right hippocampus were removed and then immersed in liquid nitrogen and stored at -70°C until ready for NOS activity measurement. For inhibition of enzyme protease all separated hippocampus (in a group) homogenized for 15 min in a solution that containing (50 mM Tris-Hcl, 0.1 mM EDTA, 0.1% 2 -me, 1 mM leopeptin, 1 mM PMSF, 2 mg L-1 aprotinin. A volume of sample containing 500 μg of protein was incubated in the presence of 10 μM L-arginin, 0.2 μCi [3H] L-arginin, 1 mM NADPH, 10 μM calmoduline and 2 mM CaCl 2. The reaction was stopped by addition of 1 mL stopping buffer that contain (3.5 mM EDTA, 20 mM HEPES). The mixture was then runs onto a resin column that equilibrated with stop buffer Results Were expressed as ng of L-Citrulline/mg protein/30 min. Because c NOS have calcium dependent activity and iNOS have calcium-inderpendent activity the enzyme activity were evaluated either in the presence of 2.5 mM and the absence of calcium in incubation medium (Diaz-Ruiz et al., 2007). Protein concentration was measured by the Lowry method (Lowry et al., 1954).
Statistical analysis: The data obtained from counting of protein concentrations. And analyzed by using SPSS version 12. Then variables have been analyzed by one-way Analysis of Variance (ANOVA) followed by Tukey post hoc test. All data were expressed as the Mean±SD and the differences were considered to be significant when p<0.05.
RESULTS
Results of NOS activities in Fig. 1 and 2 have shown that in the control groups hippocampus have cNOS (17.92±0/7) and iNOS (13.61±0/64) basal activities. Both enzymes presented a trend to increase their activity in injured animals. cNOS activity was increased (Fig. 1) in the injured animal after 2 h (28.19±0/58) (157%) and gradually increased to maximum concentration at 8 h (44.86±0/66) (250%) after injury and then slowly reduced to at 72 h (17.23±0/93). In the injured animals in the groups of rats were killed at 12, 24 and 72 h are near or equal to control levels. iNOS activities in the brain of control rats was (13.61±0/64). Interestingly, this enzyme gradually increased in injured animals.
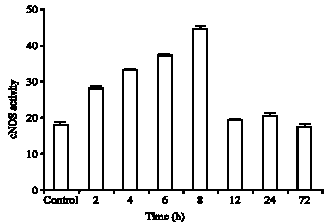 | |
| Fig. 1: | Constitutive nitric oxide synthase (cNOS) activity measured in several time after brain injury (2, 4, 6, 8, 12, 24 and 72 h). Control group received only skull drain and was scarified 2 h after injury. The results are expressed as Mean±SD (p<0.05) |
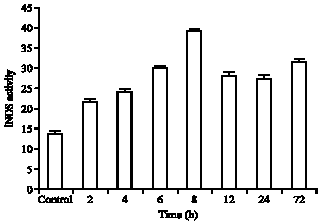 | |
| Fig. 2: | Inducible nitric oxide synthase (iNOS) activity measured in several time after brain injury (2, 4, 6, 8, 12, 24 and 72 h). Control group received only skull drain and was scarified 2 h after injury. The results are expressed as Mean±SD (p<0.05) |
After 8 h that was maximum concentration (39.11±0/50) (287%) and then slowly decreased to 72 h after injury (31.3±0/73) (230%) but are still well above control levels (Fig. 1, 2). cNOS activity was expressed higher than iNOS activity, but iNOS activity was expressed at between 8 to 72 h more significantly than cNOS activity.
Concentration of cNOS activity compared to iNOS at 8 h post injury was significant.
DISCUSSION
In the present study, the activity of both isoformes of NOS was measured at different period of time in brain injury. Results indicate a time dependent sequential increase activity in both cNOS and iNOS enzyme after injury.
Present results are agreement with data of Diaz-Ruiz et al. (2002) and Hu et al. (2008) who showed activity of cNOS increased within the first 4 h after spinal cord injury and then gradually decreased to 72 h and the maximum concentration was in 4 h after surgery, but the present data show activity of cNOS increased within the first 8 h after injury differences in the site of damage could be responsible for these versatilities.
cNOS activity after 4 h was 138% (Cherian et al., 2000) but in this study the percent of cNOS increased to 250% after 8 h and this data is considerable of course the base of cNOS in the brain is higher than spinal cord and since NO have a neuroprotective effects as a vasodilatation and neuronal function the injury of the brain cause a significant increase of NO in the brain.
Injury of the brain leads to a strong up regulation of iNOS production in a time dependent manner so cause increase iNOS protein level in injured site. According to study of Huttemann et al. (2008) the maximal level of iNOS was 4 h after injury then decreased to 48 h. But the peak activity of iNOS in our result was 8 h and then reduced. Induction of brain injury that used by Huttemann et al. (2008) cause damage to expanded area of brain and this wide damage cause earlier pick activity of iNOS.
Increased cNOS activity at short time 8 h is perhaps related to calcium dependent mechanisms evoked by the tissue response to damage (Liu et al., 1991). And Increase of iNOS activity is subsequent inflammatory response (Clark et al., 1996) and macrophages recruitment after damage (Clark et al., 1996) because the peak activity of iNOS was lower than cNOS level. Present results showed an earlier pick activity in both enzymes.
The increase in NO is subsequent higher concentration of nitrate/nitrite during the first 20 min after injury (Hu et al., 2008). Sakomoto (1997) demonstrate a nearly 2.5 fold increase in the concentration of NO in 10 min after brain injury. The mechanism of the initial increase in NO isn’t completely understood. Extraordinary metabolic changes occur in the time of brain injury and after that. At the time of injury sever depolarization occurs in the injury site and around cells, potassium concentration in extracellular space and calcium in intra cellular increase (Yamada and Nabeshima, 1997). The brain glucose increase Glutamate release. So, increase of NO based on these results is that glutamate stimulate the NMDA receptors and then causes influx of post synaptic calcium and therefore activation of the constitutive isoformes of NO.
Some research show concentration of NO reduces immediately after brain injury. The mechanism of reduce is not well understood in the present study cNOS increase 2 h after injury and then reduced 12 h post injury. Local depletion of substrate for NOS may play a role in reducing NO production (Leone et al., 1991). Polyamines ornithine decarboxylas elevated after trauma (Henley et al., 1996). The effect of this amine is cytotoxic. Polyamines are toxic to granule cells (Sparapani et al., 1997). So, effects by inhibiting NOS or by blocking inward rectifier K+ channels (Ficker et al., 1994). Free radicals such as superoxide produce after injury and may be inactivated NO and this is a mechanism for reduction of NO after 12 h post injury (Dewitt et al., 1997).
REFERENCES
- Bredt, D.S. and S.H. Synder, 1990. Isolation of nitric oxide synthase a calmodulin- requiring enzyme. Proc. Natl. Acad. Sci. USA., 87: 682-685.
PubMedDirect Link - Cherian, L., J.C. Goodman and C.S. Robertson, 2000. Brain nitric oxide changes after controlled cortical impact injury in rats. J. Physiol., 83: 2171-2178.
Direct Link - Dewitt, D.S., T.G. Smith, D.J. Deyo, K.R. Miller, T. Uchida and D.S. Prough, 1997. L-arginine and superoxide dismutase prevent or reverse cerebral hypo perfusion after fluid-percussion traumatic brain injury. J. Neurotrauma, 14: 223-233.
PubMedDirect Link - Delwing, D., D. Delwing, C.S. Bavaresco and A. Wyse, 2008. Protective effect of nitric oxide synthase inhibition or antioxidants on brain oxidative damage caused by intracerebroventricular arginine administration. Brain Res., 1193: 120-127.
PubMedDirect Link - Diaz- Ruiz, A., A. Ibarra, F.P. Severiano, G.C. Sahagun, I. Grijvalva and C. Rios, 2002. Constitutive and inducible nitric oxide synthase activates after spinal cord contusion in rats. Neurosci. Lett., 319: 129-132.
CrossRefDirect Link - Diaz-Ruiz, A., H. Salgado-Ceballos, S. Montes, V. Maldonado, L. Tristan, M. Alcaraz-Zubeldia and C. Rios, 2007. Acute alterations of Glutamate, Glutamin, GABA and other amino acids after spinal cord contusion in rats. Neurochem. Res., 32: 57-63.
CrossRefDirect Link - Dinerman, J.L., T.M. Dawson, M.J. Schell, A. Snowman and S.H. Snyder, 1994. Endothelial nitric oxide Synthase localized to hippocampal pyramidal cells: Implications for synaptic plasticity. Proc. Natl. Acad. Sci. USA., 91: 4214-4218.
PubMedDirect Link - Ficker, E., M. Taglialatela, B.A. Wible, C.M. Henley and A.M. Brown, 1994. Spermin and spermidine as gating molecules for inward rectifier K+ channels. Science, 266: 1068-1072.
CrossRefDirect Link - Gham, C., S. Holmins and T. Mathiesen, 2002. Nitric oxide synthase expression after human brain contusion. Neurosergery, 50: 1319-1326.
PubMedDirect Link - Henley, C.M., C. Muszynski, L. Cherian and C.S. Robertson, 1996. Activation of ornithine decarboxylase and accumulation of putrescine after traumatic brain injury. J. Nerotrauma, 13: 487-496.
PubMedDirect Link - Hierholzer, C., B. Herbrecht, J. Menezes, J. Kane and J. MacMicking et al., 1998. Essential role of induced nitric oxide in the initiation of the inflammatory response after hemorrhagic shock. J. Exp. Med., 187: 917-928.
Direct Link - Huttemann, M., I. Lee, C.W. Kreipke and T. Petrov, 2008. Suppretion of the inducible form of nitric oxide synthase prior to traumatic brain injury improves cytochrome oxidase activity and normalizes cellular energy levels. Neuroscience, 151: 148-154.
CrossRefDirect Link - Hu, W.H., F. Li, W. Qiang, H.Y. Wang and G.Q. Wang et al., 1992. Administration of excitatory amino acid antagonists via micro dialysis attenuates the increase in glucose utilization seen following concussive brain injury. J. Cereb. Blood Flow Metab., 12: 12-24.
PubMedDirect Link - Leone, A.M., R.M., Palmer, R.G. Knowles, P.L. Francis, D.S. Ashton and S. Moncada, 1991. Constitutive and inducible nitric oxide synthases incorporate molecular oxygen into both nitric oxide and citrulline. J. Biol. Chem., 266: 23790-23795.
Direct Link - Liu, D., W. Thangnipon and D.J. Mc Adoo, 1991. Excitatory amino acid rise to toxic levels upon impact injury to rat spinal cord. Brain Res., 547: 344-348.
PubMedDirect Link - Lowry, O.H., H.T. Graham, F.B. Harris, M.K. Priebat, A.R. Marks and R.U. Bragma, 1954. The chemical measurement of histamine in blood plasma and cells. J. Pharmacol. Exp. Ther., 112: 116-126.
PubMedDirect Link - Martinez-Murillo, R., A.P. Fernandez, J. Serrano, J. Rodrigo, E. Salas, M. Mourelle and A. Martinez, 2007. The nitric oxide donor LA 419 decreases brain damage in the focal ischemia model. Neurosci. Lett., 415: 149-153.
CrossRefDirect Link - Matsumoto, T., J.S. Pollock, M. Nakane and U. Forstermann, 1993. Developmental changes of cytosolic and particulate nitric oxide synthase in rat brain. Brain Res. Dev. Brain Res., 73: 199-203.
PubMedDirect Link - Sakomoto, K.I., H. Fujisawa, H. Koizomi, E. Tsuchida, H. Ito, D. Sadamitsu and T. Maekawa, 1997. Effects of mild hypothermia on nitric oxide synthesis following contusion trauma in the rat. J. Neurotrauma, 14: 349-353.
PubMedDirect Link - Sardo, P., G.D. Ferraro, G. Giovanni, S. Galati and V. La Grutta, 2002. Inhibition of nitric oxide synthase influences the activity of striatal neurons in the rat. Neurosci. Lett., 325: 179-182.
CrossRefDirect Link - Sparapani, M., R.O. Dall, O. Gandolfi, E. Ciani and A. Contestabile, 1997. Neurotoxicity of polyamines and pharmacological neuroprotection in culture of rat cerebellar granule cells. Exp. Neurol., 148: 157-166.
CrossRefDirect Link - Xu, M., Y.K. Ng and S.K. Leong, 2000. Neuroprotective and neurodestructive function of nitric oxide after spinal cord hemisection. Exp. Neurol., 161: 472-480.
CrossRefDirect Link - Yamada, K. and T. Nabeshima, 1997. Simultaneous measurement of nitric and nitrate levels and indices of nitric oxide release in cerebellum of conscious rats. J. Neurochem., 68: 1234-1243.
Direct Link