
Pitipong Thobunluepop
Department of Agricultural Technology, Faculty of Technology, Mahasarakham University, Talard Sub-District, Muang District, Maha Sarakham 44000, Thailand
Pakistan Journal of Biological Sciences
Year: 2009 | Volume: 12 | Issue: 16 | Page No.: 1119-1126







ABSTRACT
This study was aimed to evaluate the several chemical compounds of relatively composite structure with antifungal activity from Thai local medical plants. The antifungal activity of Stemona curtisii HK. f., Stemona tuberose L., Acorus calamus L., Eugenia caryophyllus, Memmea siamensis Kost. and an eugenol active compound were studied in vitro. Four pathogenic seed borne fungi, Alternaria solani, Colletotrichum sp., Fusarium moniliforme and Rhizoctonia solani were used as target organisms. The agar overlay technique and spore inhibition techniques were applied for the determination of their essential oil and active compound antifungal activity at various concentration; 0.10, 0.25, 0.50 and 1.00% (v/v) and untreated as control (0% v/v). Eugenol active compound showed the strongest antifungal activity on all species of tested fungal species. On the other hand, the antifungal activity of those bio-fungicides was lined up into a series from strong to low, as follows: Eugenia caryophyllus>Acorus calamus Linn.>Stemona tuberosa L.>Stemona curtisii Hk.f, while Mammea siamensis Kost. could not control any fungal species. Moreover, after eugenol application, lysis of spore and inhibition of mycelium growth were detected. Microscopic analysis exhibited complete lysis of spores after 24 h at a concentration of 1.00% v/v. Moreover, at the same concentration and 96 h incubation the mycelia growth was completely inhibited.
PDF Abstract XML References Citation
How to cite this article
Pitipong Thobunluepop, 2009. Implementation of Bio-Fungicides and Seed Treatment in Organic Rice cv. KDML 105 Farming. Pakistan Journal of Biological Sciences, 12: 1119-1126.
DOI: 10.3923/pjbs.2009.1119.1126
URL: https://scialert.net/abstract/?doi=pjbs.2009.1119.1126
DOI: 10.3923/pjbs.2009.1119.1126
URL: https://scialert.net/abstract/?doi=pjbs.2009.1119.1126
INTRODUCTION
Rice production is known to be attacked by many pathogenic fungi, are Alternaria sp., Colletotrichum sp., Fusarium sp. and Rhizoctonia sp. These microorganisms cause a serious deterioration when they occur in/on rice grain. Moreover, they can colonize to diverse substrates, because of their powerful arsenal of hydrolytic enzymes and they can be responsible for considerable economic losses (Zhu, 1998). Furthermore, some of the above-mentioned fungi can act as potential producer of mycotoxins, which potentially damage consumers’ health (Evandro et al., 2005).
Traditionally, chemical treatments are used widely to protect the germinating seedling, during vegetative and reproductive growth and after harvest from pathogenic fungi infection (Aleieri et al., 1984). Currently, the use and expectations of chemical treatments are greatly concerned due to the impact of the chemical to the environment, which can be primary or secondary influences on human or other living organisms (Baruah et al., 1996). To avoid these disadvantages, new strategies for fungicide use and disease management must be developed and identified. The alternatives of synthetic fungicides could be the development of effective phyto-chemicals from plant origin, which are expected to be more advantageous than synthetic fungicides (Davidson and Parish, 1989). The increased importance of the development and application of biological fungicides is recognized under the concept of Integrated Pest Management (IPM) (Del-Campo et al., 2002). Under this concept, all possible modes of plant disease control methods should be integrated to minimize the excessive use of synthetic fungicides (Bishop and Thornton, 1997).
It is well established that some plants contain active compounds which are able to inhibit the microbial growth (Naqui et al., 1994). These plant compounds have different structures and antimicrobial activities when compared with conventional fungicides (Nascimento et al., 2000). The potential antimicrobial properties of plant is related to their ability to synthesize several chemical compounds of relatively complex structure with antimicrobial activity, including alkaloids, flavanoids, isoflavanoids, tannins, cumanins, glycosides, terpenes and organic acids (Nychas, 1996). For examples, a solvent extract from clove flower buds that contains eugenol as main active compound was antifungal active against Alternaria sp., Fusarium sp., Botrytis sp. and Rhizoctonia sp. (Soatthiamroong et al., 2003). Sage oil was active against Botrytis sp. (Carta et al., 1996) and thyme inhibited post harvest diseases of tomato (Plotto et al., 2003).
Thus, the aim of this study was to screen for the best in vitro antifungal activity of Acorus calamus L., Stemona curtisii HK.f., Stemona tuberosa L., Memmea siamensis Kost. and Eugenia caryophyllus (Spreng.) Bullock and S.G. Harrisoncrude extracts and eugenol essential oil against rice pathogenic fungi as a possible alternative for synthetic chemical antifungal compounds.
MATERIALS AND METHODS
The experiment was conducted at Department of Agricultural Technology, Faculty of Technology, Maha Sarakham University, Maha Sarakham, Thailand and Seed Science and Technology Laboratory, Section of Seed Science and Technology, Department of Agronomy, Faculty of Agriculture, Chiang Mai University, Thailand in 2008. Plant extracts and an essential oil were used as bio-fungicides. Acorus calamus Linn., Stemona curtisii Hk.f., Stemona tuberosa L., Memmea siamensis Kost. and Eugenia caryophyllus were stream extracted and eugenol essential oil was purchased from Fluka (Steinheim, Germany). The experiment was conducted in Factorial in CRD design with 4 replications. The mycelium growth and spore germination inhibition techniques were applied to record the efficiency of those essential oils at 0.10, 0.25, 0.50 and 1.00% (v/v) and un-used of essential oils were subjected as control. Alternaria solani, Aspergillus flavus, Aspergillus niger, Bipolaris oryzae, Colletotrichum sp., Fusarium moniliforme, Nigrospora sp. and Rhizoctonia solani were used as target fungi which was provided from the collection of the Department of Plant Pathology, Faculty of Agriculture, Chiang Mai University, Thailand.
Mycelium growth inhibition analysis: The experiments were conducted by agar overlay technique was described by Morris et al. (1979). The plant extract and essential oil at different concentrations (0.10, 0.25, 0.50 and 1.00% v/v) on Potato Dextrose Agar (PDA) medium was in vitro tested against the fungi mycelium growth. Medium (20 mL) was dispensed into Petri dish and 5 mm diameter of the test fungi cutted from the middle of 7-days-old cultures were incubated upside down separately to each assay plate and incubated for 96 h at 25±2°C. The colony diameter was measured and the mycelium inhibition percentage was calculated by following Deans and Svoboda (1990). Four replicates of each treatment were tested and the average was calculated. Control sets were simultaneously run without using the plant extract and essential oil and mycelium growth inhibition was calculated as Eq. 1.
Spores inhibition analysis: Spores of Alternaria solani, Fusarium moniliforme and Rhizoctonia solani were produced by using a modification method of Mitchell and Yang (1966). The fungus was grown for 3 days at 25±2°C in PDA aqueous medium. To induce spores formation, fungal mycelium was rinsed 3 times with distilled water, for approximately 30 sec each time and after the final rinsed, cultures were kept at 25±2°C for 2 days. Spore concentration was adjusted to approximately to 106 cfu mL-1 by the hemacytometer. Sterile microscope slides were dropped 100 μL of PDA aqueous medium to obtain a thin agar layer on the slide, then 100 μL of spores suspension sample was gently spread on each slide. An uncovered watch glass containing either 100 μL of sterile water as control, or 100 μL of plant extract and an essential oil at different concentration (0.10, 0.25, 0.50 and 1.00% v/v) were dropped into slide. After that, slides were place in glass Petri dishes lined with moistened filter paper, covered and sealed with parafilm, incubated for 24 h at 25±2°C, all of the encysted spores on each slide were counted with compound microscope at magnification of x100 and spores inhibition was calculated as Eq. 1.
Inhibition (%) =[(C-T)/C] × 100(1) | (1) |
where, C is the colony diameter of the mycelium on the control plate (mm) and T is the colony diameter of the mycelium on the treatment plate (mm).
Concentration response curves were obtained whereby the percentage of fungal inhibition was plotted against concentration and concentration required to give 50% inhibition of fungal growth (IC50) was calculated from the regression equation.
Statistical analysis: The data are presented as Mean±Standard deviation. The analysis of variance was performed for data analysis and differentiated with Last Significant Different (LSD) test at p<0.05 using the software SX release 8.0 (Analytical software, Tallahassee, USA).
RESULTS AND DISCUSSION
The most of studied fungi were inhibited at 1% v/v of A. calamus Linn.; A. solani, Colletotrichum sp. and R. solani were completely inhibited (100%). Nevertheless, F. moniliforme was inhibited about 95.91% (Fig. 1). R. solani was slightly sensitive to S. curtisii Hk.f. extract and was inhibited to 49.54% at 1.00% v/v concentration.
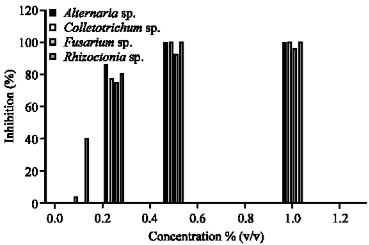 | |
| Fig: 1 | The antifungal activity of Acorus calamus L. against economically rice pathogenic fungi |
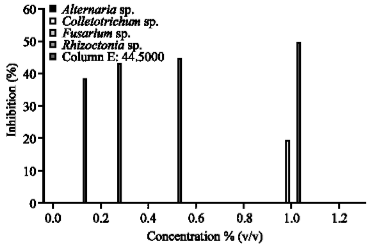 | |
| Fig. 2: | The antifungal activity of Stemona curtisii Hk.f. against economically rice pathogenic fungi |
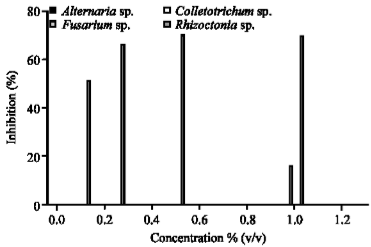 | |
| Fig. 3: | The antifungal activity of Stemona tuberosa L. against economically rice pathogenic fungi |
The growth of A. solani, Colletotrichum sp. and F. moniliforme were uninhibited by S. curtisii Hk.f. extract (Fig. 2). Figure 3 shows the antifungal activity of S. tuberosa L. which had no inhibition effect on A. solani, Colletotrichum sp. and F. moniliforme. However, S. tuberosa L. could inhibited R. solani for 70% at 1.00% v/v concentration.
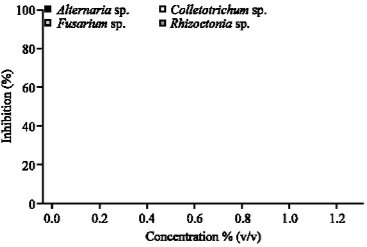 | |
| Fig. 4: | The antifungal activity of Memmea siamensis Kost. against economically rice pathogenic fungi |
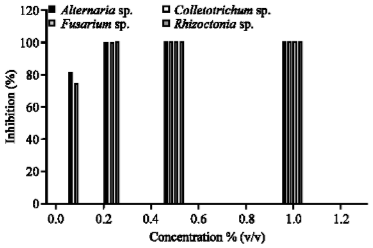 | |
| Fig. 5: | The antifungal activity of Eugenia caryophyllus against economically rice pathogenic fungi |
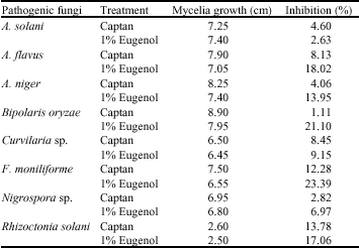
As shown in Fig. 4, the M. siamensis Kost extract was unable to control all fungi species. Figure 5 showed that E. caryophyllus extract at 0.25% v/v completely inhibited A. solani, Colletotrichum sp. and F. moniliforme. Moreover, when the concentration increased to 0.50% v/v, this extract inhibited Rhizoctonia solani completely. The IC50 values indicated that E. caryophyllus crude extract showed strongly antifungal activity against pathogenic fungal as well as A. calamus Lin. However, S. curtisii Hk.f, S. tuberosa L. and M. siamensis Kost. were unable to inhibit all fungi species. Although, R. solani seemed to be less susceptible to the plant extracts than the other fungi (Table 1). Table 2 shows that, eugenol showed the strongest antifungal activity at 1.00% v/v. Additionally, eugenol had the strongest antifungal activity (showed 100% inhibition zone) against the each fungal species as well as E. caryophyllus did (Table 3). eugenol was similar effective in inhibiting the mycelium growth of each fungus as captan did, especially on A. solani, A. flavus, A. niger, B. oryzae and F. moniliforme (Table 4, Fig. 6). Nevertheless, captan was effective on inhibition of spore germination completely at 0.10% v/v but eugenol significantly inhibited fungal spore germination only at 0.50 or 1.00% v/v (Table 5, Fig. 7).
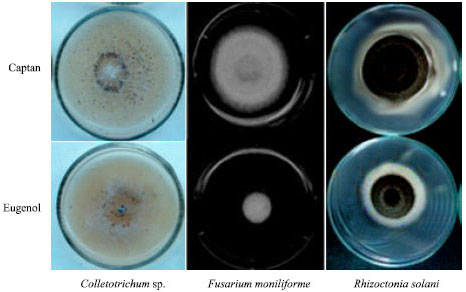 | |
| Fig: 6 | Comparative the antifungal activity of synthetic fungicide; Captan and eugenol botanical fungicides; eugenol treated at 1.00% v/v against fungal mycelium growth inhibition |
| Table 1: | The 50% inhibitory concentration (IC50) of botanical fungicides against some species of rice pathogenic fungi |
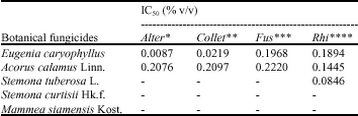 | |
| *Alternaria solani, **Colletotrichum sp., ***Fusarium moniliforme, ****Rhizoctonia solani. IC50: The concentration required to give 50% inhibition of fungal growth, - : Not inhibited | |
| Table 2: | The concentration of botanical fungicide; eugenol against rice pathogenic fungi |
 | |
| Table 3: | Comparative the antifungal activity of Eugenia caryophyllus crude extract and eugenol (purified) active compound against rice pathogenic fungi |
 | |
| *Alternaria solani, **Colletotrichum sp., ***Fusarium moniliforme, ****Rhizoctonia solani | |
| Table 4: | Comparative the antifungal activity of synthetic fungicide; captan and botanical fungicide; eugenol treatment on fungal mycelia growth inhibition |
 | |
The antifungal activity of plant crude extracts, or essential oil against pathogenic fungi can be lined up in order from strongest to lowest effectiveness as follows: eugenol>Eugenia caryophyllus>Acorus calamus Linn.>Stemona tuberosa L.> Mammea siamensis Kost.= Stemona curtisii Hk.f.
At low concentration, S. tuberosa L. and S. curtisii Hk.f. crude extracts were unable to control all species of fungi. Although, at the highest assayed concentration (1.0% v/v), S. tuberose L. extract inhibited only Rhizoctonia sp. The previous studies reported that S. curtisii Hk.f. and S. tuberose L. crude extracts had much stronger insecticidal activity than fungicidal activity (Chantawannakul et al., 2005). Furthermore, Issakul et al. (2004) found that A. calamus Linn. extract showed strong antifungal activity but the M. siamensis Kost. crude extract has no antifungal activity.
 | |
| Fig. 7: | The antifungal activity at various concentrations of botanical fungicide; eugenol treatment against Fusarium moniliforme spores germination, studies showed that eugenol at concentration higher than 0.5% (v/v) can completely inhibit the spore germination |
| Table 5: | Comparative the effect of Captan, Eugenia caryophyllus and eugenol treatment on the inhibition of spore germination of some species of rice pathogenic fungi |
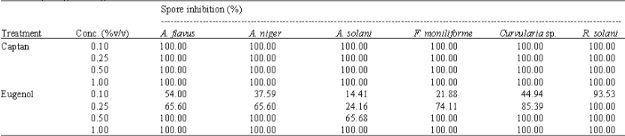 | |
However, M. siamensis Kost was previous found to have a strong insecticidal activity.
The present study found that A. calamus Linn. (Sweet flag) has an effective antifungal activity, may be ²μ-asarone and a-asarone are the major constituents in the rhizomes and leaf oils of A. calamus Linn (Raina et al., 2003). These compounds are provided antifungal active against Curvularia sp. and Alternaria sp. (Janssen et al., 1988). Ghosh (2006) reported the A. calamus Linn. crude extract inhibited hyphal growth of F. moniliforme, whereas, Lee (2006) found that methanolic extract of A. calamus L. had strong fungicidal activity against R. solani. Nevertheless, the results suggested that E. caryophyllus crude extract had the strongest antifungal activity at its lowest concentration.
The concentration and active compound of plant products are the most important factors in the antifungal activity. Obviously, the results indicated that at high concentration (1.00% v/v) of E. caryophyllus crude extract could inhibit the pathogenic fungi. However, in term of spore germination and mycelia growth inhibition, eugenol was found to have stronger inhibition ability than E. caryophyllus crude extract, which was in agreement with Viollon and Chaumont (1994). eugenol is the major phenolic compound of E. caryophyllus and have a strong antibacterial and antifungal activity, e.g., against Aspergillus sp. and Fusarium sp. (Pauli and Knobloch, 1987), as well as against Alternaria sp., Fusarium sp., Curvularia sp. and Rhizoctonia sp. (Beg and Ahmed, 2002). The present study established the high effectiveness of eugenol at the Minimum Fungistatic Concentration (MFC) of 0.50% v/v. At concentrations higher than 0.50% v/v, lysis of spore and inhibition of mycelia growth were detected and confirm the results of Neni et al. (2006). However, the MFC and toxicity concentration varied from study to study. This is probably due to the different extraction methods of essential oils and different sensitivity of the test strains used (Saikia et al., 2001). The plant extracts showed clear antimicrobial properties, although the mechanistic of action are poorly understood. However, it must be pointed out that the intrinsic activity of a compound is very important for its effectiveness. In this context, the essential oil containing phenolic compound was reported to exhibit a high inhibitory effect (Bennis et al., 2004).
The mode of action of antifungal agents depends also on the type of target microorganisms and is mainly related to their cell wall structure and the outer membrane arrangement (Dorman and Deans, 2000). These observations suggested that the physical and chemical properties (solubility and volatility) might have considerably effects on the in vitro antimicrobial activity of plant extracts (Inouye et al., 2000). eugenol was found in this study to strong activity due to their relatively low capacity to dissolve in water, which supported by Hilli et al. (1997). The effectiveness of eugenol depends on the structure of the phenolics where in previous studies identified as active compounds, e.g., the hydroxyl group and its relative position within the molecule (Tullio et al., 2006). High hydrophobic compounds are generally reported to be very effective on the primary site at the cytoplasmic membrane (Sikkema et al., 1995). The effect of eugenol when separated from the fungi membranes suggests that its activity is based on the lipophilic properties. The interactions between antimicrobial compounds and cell membranes affect both the lipid ordering and the bilayer stability (Ben-Arfa et al., 2006). Their mode of action appeared to be at the phospholipid bilayer, caused by biochemical mechanisms, catalysed by the phospholipid bilayers of the cell and related to the cell membrane disruption. These processes include the inhibition of electron transport, protein translocation, phosphorylation steps and other enzyme-dependent reaction (Knobloch et al., 1988). eugenol is able to inhibit the respiration and ion transport processes, increase membrane permeability and the releasing of cellular content (Uribe et al., 1985). Moreover, eugenol is able to inhibit the respiration of cell suspensions and to disrupt the permeability barrier of microbial membrane structure (Cox et al., 2000).
Morris et al. (1979) reported that the fungi after treated with eugenol, decreased its size, appeared in irregular shape with cell wall modifications and the cell surface depressions. Such modifications may be related to the interference of the oil components with enzymatic reactions of cell wall synthesis, which affects fungal morphogenesis and inhibited growth.
According to Helal et al. (2006), the antifungal activity of eugenol may take place via two steps. The first step involves the passive entry of the oil into the plasma membrane in order to initiate cell membrane disruption. The second step, the accumulation of oil in the plasma membrane results in the inhibition of cell growth. This can be ascribed as combination of membrane effects such as increased bi-layer disorder and ion leakage. These effects disturb the osmotic balance of the cell through the loss of ions, making its membrane associated proteins inefficient due to increased membrane disorder eventually leading to inhibition of cell growth. The cytoplasmic, plasma and mitochodrial membrane of fungal provide a barrier to the passage of small ion such as H+, K+, Na+ and Ca2+ and allow cells and organelles to control the entry and exit of different compounds (Suhr and Nieken, 2003). This permeability barrier role of cell membranes is integral to many cellular functions, including the maintenance of the energy status of the cell, other membrane-coupled energy-transducing process, solute transportation, regulation of metabolism and control of turgor pressure (Trumpower and Gennis, 1994). Cox et al. (2000) observed the leakage of the K+, Ca2+ and Mg2+ from exposed fungal cells with eugenol. Some changes in the cell membrane may occur in spite of the damage to the plasma membrane. These changes are accompanied with the loss of chemiosmotic control disrupted the permeability barrier of cell membrane structure. Ultee et al. (2002) hypothesized that the hydroxyl group and the presence of a system of delocalized electrons are important for the antimicrobial activity of the eugenol. Such a particular structure would allow compounds to act as proton exchanger, thereby reducing the pH gradient across the cytoplasmic membrane. Eventually, the reduction of the proton motive force and the depletion of the ATP pool lead to the cell death. Hence, these findings supported our results that eugenol was identified as the best antifungal effect of all studied plant extracts.
CONCLUSION
The crude plant extracts from Stemona curtisii Hk.f., Stemona tuberosa L. and Mammea siamensis Kost. did not have an antifungal activity. However, high concentrations of Acorus calamus Linn. and the Eugenia caryophyllus crude extracts can inhibit the studied pathogenic fungi. eugenol, an active compound of Eugenia caryophyllus, showed the strongest antifungal activity in inhibiting the growth of pathogenic fungi. The mode of action based on the deterioration of the fungal cellular structure leading to complete cell death, even at lower concentration of eugenol. Therefore, based on the antifungal activity against pathogenic fungi, the following order from strongest to lowest effectiveness was found: eugenol > Eugenia caryophyllus > Acorus calamus Linn. > Stemona tuberosa L. > Mammea siamensis Kost. = Stemona curtisii Hk.f. This finding increases the possibility of exploiting eugenol as a promising candidate for safe natural antifungal agent.
ACKNOWLEDGMENTS
Authors acknowledge the Nation Research Council of Thailand (NRCT), Royal Thai Government, for research financial supported of the present studies, gratefully thanks to Dr. Suchada Vearasilp, Prof. Dr. Elke Pawelzik, Prof. Dr. Udo ter Meullen, Prof. Dr. Wolfgang Lüke for theirs are valuable suggestions through out the research project.
REFERENCES
- Baruah, P., R.K. Sharma, R.S. Singh and A.C. Ghosh, 1996. Fungicidal activity of some naturally occurring essential oils against Fusarium moniliforme. J. Essential Oil Res., 8: 411-412.
CrossRefDirect Link - ben Arfa, A., S. Combes, L. Preziosi-Belloy, N. Gontard and P. Chalier, 2006. Antimicrobial activity of carvacrol related to its chemical structure. Lett. Appl. Microbiol., 43: 149-154.
CrossRefDirect Link - Bennis, S., F. Chami, N. Chami, T. Bouchikhi and A. Remmal, 2004. Surface alteration of Saccharomyces cerevisiae induced by thymol and eugenol. Lett. Appl. Microbiol., 38: 454-458.
CrossRef - Bishop, C.D. and I.B. Thornton, 1997. Evaluation of the antifungal activity of the essential oils of Monarda citriodora var. citriodora and Melaleuca alternifolia on post-harvest pathogens. J. Essential Oil Res., 9: 77-82.
CrossRefDirect Link - Cox, S.D., C.M. Mann, J.L. Markham, H.C. Bell, J.E. Gustafson, J.R. Warmington and S.G. Wyllie, 2000. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Applied Microbiol., 88: 170-175.
CrossRefPubMedDirect Link - Davidson, P.M. and M.E. Parish, 1989. Methods for testing the efficacy of food antimicrobials. Food Technol., 43: 148-155.
Direct Link - Deans, S.G. and K.P. Svoboda, 1990. The antifungal properties of margoram (Origanum majorana L.) volatile oil. Flavour Fragrance J., 5: 187-190.
Direct Link - Dorman, H.J.D. and S.G. Deans, 2000. Antimicrobial agents from plants: Antibacterial activity of plant volatile oils. J. Appl. Microbiol., 88: 308-316.
CrossRefDirect Link - Ghosh, M., 2006. Antifungal properties of Haem peroxidase from Acorus calamus. Ann. Bot., 98: 1145-1153.
CrossRefDirect Link - Helal, G.A., M.M. Sarhan, A.N.K. Abu-Shahla and E.K. Abou-El-Khair, 2006. Effect of Cymbopogon citrates L. essential oil on growth and morphogenesis of Saccharomyces cerevisiae ML2-strain. J. Basic Microbiol., 46: 375-386.
CrossRefDirect Link - Hili, P., C.S. Evans and R.G. Veness, 1997. Antimicrobial action of essential oils : The effect of dimethylsulphoxide on the activity of cinnamon oil. Lett. Applied Microbiol., 21: 269-275.
CrossRefDirect Link - Issakul, K., W. Kongtrakoon, S. Dheeranupatan, S. Jangsutthivoranat and A. Jatisatienr, 2004. Insecticidal effect of compounds from Mammea siamensis Kost. against Musca domestica Linn. Acta Hortc., 629: 103-110.
Direct Link - Morris, J.A., A. Khettry and E.W. Seitz, 1979. Antimicrobial activity of aroma chemicals and essential oils. J. Am. Oil Chem. Soc., 56: 595-603.
CrossRef - Neri, F., M. Mari and S. Brigati, 2006. Control of Penicillium expansum by plant volatile compounds. Plant Pathol., 55: 100-105.
CrossRefDirect Link - Pauli, A. and K. Knobloch, 1987. Inhibitory effect of essential oils components on growth of food-contaminating fungi. Z Lebensm Unters Forsch, 185: 10-13.
CrossRef - Plotto, A., D.D. Roberts and R.G. Roberts, 2003. Evaluation of plant essential oils as natural postharvest disease control of tomato (Lycopersicon esculentum). Acta Hortic., 628: 737-745.
Direct Link - Raina, V.K., S.K. Srivastava and K.V. Syamasunder, 2003. Essential oil composition of Acorus calamus L. from the lower region of the Himalayas. Flavour Fragrance J., 18: 18-20.
CrossRefDirect Link - Saikia, D., S.P.S. Khanuja, A.P. Kahol, S.C. Gupta and S. Kumar, 2001. Comparative antifungal activity of essential oils and constituents from three distinct genotypes of Cymbopogon spp. Curr. Sci., 86: 1264-1266.
Direct Link - Sikkema, J., J.A. de Bont and B. Poolman, 1995. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Mol. Biol. Rev., 59: 201-222.
PubMedDirect Link - Suhr, K.I. and P.V. Nieken, 2003. Antifungal activity of essential oils evaluated by two different application techniques against rye bread spoilage fungi. J. Applied Microbiol., 94: 665-674.
CrossRef - Trumpower, B.L. and R.B. Gennis, 1994. Energy transduction by cytochrome complexes in mitochondrial and bacterial respiration: The enzymology of coupling electron transfer reactions to trans-membrane proton translocation. Annu. Rev. Biochem., 36: 675-716.
Direct Link - Uribe, S., T. Samirez and A. Pena, 1985. Effect of β-pinene on yeast membrane functions. J. Bacteriol, 161: 1195-1200.
Direct Link - Viollon, C. and J.P. Chaumont, 1994. Antifungal properties of essential oils and their main components upon Cryptococcus neoformans. Mycopathologia, 128: 151-153.
CrossRef - Beg, Z.A. and I. Ahmed, 2002. In vitro fungitoxicity of the essential oil of Syzygium aromaticum. World J. Microbiol. Biotechnol., 18: 317-319.
Direct Link - Janssen, A.M., J.J.C. Scheffer, A.W.P. van Atte and A.B. Svendsen, 1988. Screening of some essential oil for their activities on dermatophytes. Pharm. Weekbl. Sci., 10: 277-280.
CrossRefPubMedDirect Link - Tullio V., A. Nostro, N. Mandras, P. Dugo and G. Banche et al., 2006. Antifungal activity of essential oils against filamentous fungi determined by broth microdilution and vapor contact methods. J. Applied Microbiol., 102: 1544-1550.
CrossRefDirect Link - Ultee, A., M.H.J. Bennik and R. Moezelaar, 2002. The phenolic hydroxyl group of Carvacrol is essential for action against the food-borne pathogen Bacillus cereus. Applied Environ. Microbiol., 68: 1561-1568.
CrossRefDirect Link - De Souza, E.L., E. de Oliveira Lima, K.R. de Luna Freire and C.P. de Sousa, 2005. Inhibitory action of some essential oils and phytochemicals on the growth of various moulds isolated from foods. Braz. Arch. Biol. Technol., 48: 245-250.
CrossRefDirect Link