
Yahya Kooch
Department of Forestry, Faculty of Natural Resources,University of Agriculture and Natural Resources of Sari, Islamic Republic of Iran
Hamid Jalilvand
Department of Forestry, Faculty of Natural Resources,University of Agriculture and Natural Resources of Sari, Islamic Republic of Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 6 | Page No.: 819-825







ABSTRACT
Earthworms are considered as soil engineers because of their effects on soil properties and their influence on the availability of resources for other organisms, including microorganisms and plants. However, the links between their impacts on the soil environment and the resulting modification of natural selection pressures on engineer as well as on other organisms have received little attention. Earthworms are known to have a positive influence on the soil fabric and on the decomposition and mineralization of litter by breaking down organic matter and producing large amounts of fasces, thereby mixing litter with the mineral soil. Therefore, they play an important part in changes from one humus from to another according to forest succession patterns. Consequently, they are also expected to be good bio-indicators for forest site quality and are thus useful when planning forest production improvement. Earthworm`s populations are as indicator that in exploited regions is destruction indicator and reclamation plans is nature return indicator. In this study we summarized the current knowledge in relation to earthworm`s ecology in forest soils as ecosystem engineers.
PDF Abstract XML References Citation
How to cite this article
Yahya Kooch and Hamid Jalilvand, 2008. Earthworms as Ecosystem Engineers and the Most Important Detritivors in Forest Soils. Pakistan Journal of Biological Sciences, 11: 819-825.
DOI: 10.3923/pjbs.2008.819.825
URL: https://scialert.net/abstract/?doi=pjbs.2008.819.825
DOI: 10.3923/pjbs.2008.819.825
URL: https://scialert.net/abstract/?doi=pjbs.2008.819.825
INTRODUCTION
Ecosystem engineering is organisms that directly or indirectly modulate the availability of resources to other species, by causing physical state changes in biotic or abiotic materials (Alban, 1982; Armson, 1979; Bohlen, 2002; Hairiah et al., 2006). Ammer et al. (2005) argues that in soil, the relative importance of regulation imposed by ecosystem engineering is likely to be greater than regulation by trophic relationships because of the specific ecological constraints observed in this environment when compared to above-ground conditions.
Earthworms, termites and ants have been identified as the most important soil engineering (Gonzalez et al., 2003). At the heart of the soil engineering concept is the ability of theses organisms to move through the soil and to build organo-mineral structures with specific physical, chemical and microbiological properties. Usually called biogenic structures; these structures can be galleries, casts, mounds, fungus comb chambers and so on (Amador et al., 2006).
Earthworms have been described as being one of the main groups of soil engineers in tropical and temperate ecosystems because they change the structural properties of soil and thus influence soil microorganisms and plant growth (Kimmins, 1987; Jongman et al., 1995; Judas, 2002; Muratake, 2005). Earthworms are perhaps the most important soil organisms in terms of their influence on organic matter breakdown, soil structural development and nutrient cycling, especially in productive ecosystems (Kooch et al., 2007).
Aristole called them the intestines of the earth and the eminent nineteenth century biologist, Charles Darwin, spent many years observing their major influence on the formation of humus and transport of soil (Johnson-Maynard et al., 2002). Despite the vast increase in scientific literature on earthworms in recent years, much remains to be known of their basic biology and ecology (Jimens et al., 2006).
Earthworms are the best known and perhaps the most important animals that live in soil. Over 3500 earthworm species have been recognized in worldwide and it is estimated that further surveys will reveal this number to be much larger (Deleporte, 2001; Hendrix and Bohlen, 2002). Earthworms alter soil properties in ways that are beneficial to plant growth by improving soil structure for better aeration, water intake and water transmission and, are known to have various beneficial effects on soil physical properties (Kimmins, 1987; Haynes et al., 2003; Rombke et al., 2005; Sautter et al., 2006).
Earthworms play a major role in soil nutrient dynamics by altering the soil physical, chemical and biological properties. Their casts, burrows and associated middens constitute a very favorable microenvironment for microbial activity (Hale et al., 2005; Hale and Host,2005). They affect nutrient cycling by modifying soil porosity (Ammer et al., 2005) and aggregates structure (Sheehan et al., 2006), changing the distribution and rates of decomposition of plant litter and altering the composition, biomass and activity of soil microbial communities (Jimens et al., 2006).
EARTHWORM BIOLOGY AND CLASSIFICATION
Earthworms are terrestrial annelids with bilateral symmetry and corresponding external and internal segmentation. They have a thinly pigmented cuticle bearing setae on all segments except the first two. All earthworms are hermaphroditic, which very among different taxa. When sexually mature, they develop a swollen area of the epidermis called a clitellum. This region produces a cocoon in which one or more eggs are deposited and then the cocoon is passed over the anterior segments and deposited in or on the soil. The young develop within the cocoon and newly hatched worms resemble adults. The time to hatching and reproductive maturity varies widely among different earthworm species and is influenced by environmental factors (Beyer et al., 1991; Binkley and Valentine, 1991; Curry, 1998; Edwards and Bohlen, 1996; Holscher et al., 1999; Grant and Loneragan, 2001; Parkinson and Welke, 2003; Ratty and Hutha, 2004).
Earthworms are classified within the phylum Annelida and the class Oligochaeta, which consists of as many as 36 families worldwide. About two-thirds of Oligochaeta families comprise aquatic or semi-aquatic worms and the remaining families comprise mostly or exclusively terrestrial worms or earthworms. There are over 3500 known earthworm species and it is estimated that the global total may twice that number. Distinct taxonomic groups have arisen on every continent except Antarctica and some groups are now distributed throughout the world (Ammer and Makaeschin, 1994; Begon et al., 1995; Wood, 1995; Warren and Zou, 2002).
EARTHWORM ECOLOGY
Ecological groups: Earthworm species can be grouped according to behavioral, morphological or physiological adaptations that enable them to partition available resources in the soil. The three main life history strategies are termed epi, aneceic and endogeic (Table 1 and Fig. 1) (Miles, 1985; Freid et al., 1989; Habibi Kaseb, 1992; Haghparast Tanha, 1993; Hedman et al., 2000).
| Table 1: | Ecological categories, habitat, feeding and morphological characteristics of earthworms |
 | |
| Sources: Aubert et al. (2003), Ratty (2004) and Iman Nejad and Rahmani (2005) | |
 | |
| Fig. 1: | Earthworm`s representative of different ecological groups, (a) Epigeic species, such as Dendrodrilus rubidus, inhabit organic rich surface layers and feed mainly on surface organic matter, (b) Endogeic species, such as Octolasion tyrtaeum, consume more mineral soil than epigeic species and mix mineral and organic soil layers together and (c) Aneceic species, such as Lumbericus terrestris, live in deep vertical burrows, feed mainly on surface litter and incorporate litter into the soil as well as transporting mineral soil to the surface from deeper soil layers (Kooch et al., 2007) |
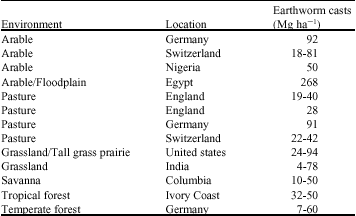
| Table 2: | Amount of organic matter ingested or incorporated into soil by earthworm populations in different environments |
 | |
| Sources: Hendrix and Bohlen (2002), Parkinson and Welke (2003), Ratty and Hutha (2004) and Sheehan et al. (2006) | |
Epigeic worms feed on plant litter, dwell on the soil surface or within the litter layer, tend to be heavily pigmented and are small to medium sized. Aneceic worms feed on plant litter and soil, live in nearly vertical permanent burrows, are dorsally pigmented and large. Endogeic species are soil-feeders, are not heavily pigmented, from extensive horizontal burrow systems and range in size form small to large. Endogeic worms have been further divided into polyhumic, mesohumic and oligohumic groups, which are separated, respectively, by the descending importance of organic rich mineral soil in their diet and increasing size. Earthworm species do not always fall clearly into these three main categories and may even exhibit traits of different groups at different life stages or under different environmental conditions (Darwin, 1881; Hurnung, 1985; Lee, 1985; Mackney, 1961; Schaller, 1968; Edwards et al., 1973; Tappeiner and Alm, 1975; Saleh Rastin, 1978; Black, 1979; Huhta, 1979; Persson, 1988; Wood and James, 1993; Norden, 1994; Raulund-Rasmussen and Vejre, 1995; Wilson and Grigal, 1995; Neirynck et al., 2000).
Earthworm communities: Earthworm communities generally consist of from one to six species. The relative abundance and species composition of earthworm communities depends upon soil type, topography and vegetation and is also influenced by land use history and earthworm biogeography (Neirynck et al., 2000; Scheu et al., 2002; Six et al., 2004).
Earthworms account for the majority of animal biomass in soil in a wide range of productive ecosystems, from temperate grasslands, pastures and forests to tropical pastures, savannas and rainforests and many temperate and tropical agro ecosystems. They generally do not occur in deserts and arid grasslands or in extreme alpine or boreal habitats. Earthworms are often absent from strongly acidic forest soils with poor litter quality, such as some northern coniferous forests (Muys and Lust, 1992; Rahmani, 1998; Scheu and Schaefer, 1998; Rahmani and Saleh Rastin, 2000; Scheu et al., 2003).
Many earthworm communities consist of invasive exotic species. In North America, where approximately 100 native earthworm species have been introduced. Lumbericid earthworms of European origin dominate many North American forest ecosystems. A worldwide survey of earthworms in tropical regions reported that a total of 51 exotic and 151 native species commonly occur in tropical forest ecosystems (Ammer et al., 2005; Hale et al., 2005; Hale and Host, 2005; Sautter et al., 2006).
Effects of earthworms on soil properties: The influence of a given earthworm species on soil properties depends upon that species life history strategy. For example, the large vertical burrows of aneceic worms, such as the common night crawler Lumbericus terrestris, can facilitate preferential flow of water through the soil profile, increasing the transport of water, nutrients and agricultural chemicals into deeper soil layers (Gonzalez et al., 2003; Hairiah et al., 2006).
Epigeic species facilitate the breakdown and mineralization of surface litter, whereas aneceic species incorporate surface litter deeper into the soil profile. Aneceic species also bring soil from deeper soil horizons to the surface, which over a long period of time can change the mineralogy of surface soil. Endogeic species feed on fragmented organic matter and mix it thoroughly through the surface mineral soil (Scheu et al., 2002; Sheehan et al., 2006).
Effects on organic matter breakdown and nutrient cycling: Earthworm activity accelerates the composition of plant litter, increase rates of nutrient transformation and plant nutrient uptake, improves soil aggregation and porosity and enhances water infiltration and solute transport. Earthworms can consume and incorporate large amounts of organic matter into soil (Table 2). Such mixing is largely responsible for the formation of mull soils in which surface organic horizons are thoroughly mixed with underlying mineral soil (Hendrix and Bohlen, 2002; Iman Nejad and Rahmani, 2005; Sautter et al., 2006).
Earthworms are major influence on nutrient cycling in many ecosystems. Although they generally increase the mineralization of soil carbon, earthworms can also decrease mineralization of carbon by contributing to the formation of stable soil aggregates in which carbon is protected from further breakdown. Direct fluxes of
| Table 3: | The amount of earthworm casts produced and in different environments in various locations around the world |
 | |
| Sources: Rahmani (1998) and Iman Nejad and Rahmani (2005) | |
nutrients through earthworm biomass can be considerable. For example, the turnover of nitrogen (N) through earthworm tissues can be up to 150 kg N ha-1 year-1 (Saleh Rastin, 1978; Six et al., 2004; Rombke et al., 2005).
Earthworm casts contain elevated amounts of inorganic N relative to surrounding soil. As a consequence, earthworms can greatly enhance the mineralization of N and can simulate other N transformation such as denitrification (Iman Nejad and Rahmani, 2005). By increasing bypass flow of infiltering water, earthworm burrows can increase the amount N and other nutrients leaching from the soil profile (Neirynck et al., 2000). Alternatively earthworms can reduce the amount of nutrients lost in surface runoff by increasing rates of water infiltration into the soil (Aubert et al., 2003).
Effects on physical properties of soil: The effect of earthworms on soil structure results from the net outcome of their feeding and burrowing activities. Earthworms ingest soil particles and organic matter, mixing these two fractions together and egesting them as surface or subsurface casts. Estimates of annual rates of production of earthworm cast range from less than 5 to over 250 Mg ha-1 in various ecosystems (Table 3) (Rahmani, 1998; Ratty, 2004; Iman Nejad and Rahmani, 2005).
Once egested, soil in casts can be eroded due to impact of rainfall or can form stable soil aggregates through in variety of stabilizing mechanisms (Saleh Rastin, 1978). Earthworms generally improve the aeration and porosity of soil thorough formation of burrows and by increasing the proportion of large aggregates in the soil and their effects are especially important in poorly structured or reclaimed soil. By increasing rates of water infiltration, earthworms can reduce the amount of surface runoff. Alternatively, earthworms can increase erosion by removing the protective cover of surface litter, increasing surface sealing and depositing surface casts, which can be carried down slope during heavy rains. Some tropical species actually increase soil bulk density and decrease infiltration by producing compact soil casts. Overall, the effects of earthworms on soil structure are considered to improve soil fertility (Aubert et al., 2003; Parkinson and Welke, 2003; Iman Nejad and Rahmani, 2005).
Effects on plant growth: The majority of studies examining the influence of earthworms on plant growth have reported that earthworms simulate plant growth, although some studies have reported no effect or even a negative effect of earthworms on plant growth (Rahmani and Saleh Rastin, 2000; Scheu et al., 2002; Judas, 2002). Earthworms have been shown to increase production of shoots and grain in a variety of field trails and greenhouse experiments.
Introduction of earthworms into reclaimed polders in the Netherlands and in pasture in New Zealand resulted in large increase in forage quantity (Hedman et al., 2000). Beneficial effects of earthworms on plant growth may be due to increased nutrient and water availability, improved soil structure, simulation of micro organisms or formation of microbial products the enhance plant growth, or possibly through direct production of plant growth promoting substances.
Undesirable effects of earthworms: Despite the documented and putative beneficial effects of earthworms on nutrient dynamics, soil structure and fertility, some aspects of earthworm activities are considered undesirable (Haynes et al., 2003). These include removing and burying surfaces from erosion; producing fresh casts that increase erosion and surface of lawns and golf greens or irrigation ditches where they are a nuisance, or in pasture where they interfere with haying operations; dispersing weed seeds in gardens and agricultural fields; transmitting plant or animal pathogens; increasing losses of soil nitrogen through leaching and denitrification and increasing soil carbon loss through enhanced microbial respiration. It is the net outcome of their positive and negative effects that determines whether earthworms are to be considered pests or benefactors in any given situation and they are generally considered to be of great benefit to soil properties and fertility (Rahmani, 1998; Rahmani and Saleh Rastin, 2000; Warren and Zou, 2002; Ratty and Hutha, 2004; Hale et al., 2005).
Earthworms and soil reclamation: Introduction of appropriate earthworm species or encouraging natural population through the addition of suitable amendments can increase the rate of soil improvement and genesis of soil structure of reclaimed land. Introduction of European earthworms into pasture in New Zealand and Australia, as well as in Dutch polders, greatly facilitated improvements in soil structure and plant productivity (Ratty, 2004). There has also been some success in introducing earthworms into reclaimed mining sites and in reclaimed peat with beneficial effects on soil structural development, nutrient cycling and productivity (Ratty and Hutha, 2004).
CONCLUSION
Earthworms are the most important members of soil detritivors in temperate forests. Soil productivity and plant growth are significantly affected by biological activities of earthworms (Rahmani, 1998; Rahmani and Saleh Rastin, 2000). Earthworms play a major role in soil nutrient dynamics by altering the soil physical, chemical and biological properties. Their casts, burrows and associated middens constitute a very favorable microenvironment for microbial activity. They affect nutrient cycling by modifying soil porosity and aggregate structure, changing the distribution and rates of decomposition of plant litter and altering the composition, biomass and activity of soil microbial communities (Hurnung, 1985; Neirynck et al., 2000; Scheu et al., 2002; Warren and Zou, 2002; Rombke et al., 2005; Sheehan et al., 2006).
Earthworm excreta, such as ammonia and urea and body tissue are rapidly mineralized, thus providing a potentially significant source of readily available nutrients for plant growth and it has been estimated that fluxes of nitrogen from earthworm population in ecosystems can range from 10 to 74 kg N/ha-1 year (Rahmani, 1998; Rahmani and Saleh Rastin, 2000; Judas, 2002; Jimens et al., 2006). Consequently, they are also expected to be good bio-indicators for forest site quality and are thus useful when planning forest production improvement. Earthworm`s populations are as indicator that in exploited regions is destruction indicator and reclamation plans is nature return indicator (Rahmani and Saleh Rastin, 2000).
REFERENCES
- Alban, D.H., 1982. Effects of nutrient accumulation by Aspen, spruce and pine on soil properties. Soil. Sci. Soc. Am. J., 46: 853-861.
CrossRef - Amador, J., J. Gorres and C. Savin, 2006. Effects of Lumbricus territories L. on nitrogen dynamics beyond the burrow. Applied Soil Econ., 33: 61-66.
Direct Link - Ammer, S., K. Weber, C. Abs, C. Ammer and J. Prietzel, 2006. Factors influencing the distribution and abundance of earthworm Communities in pure and converted scots pine stands. Applied Soil Ecology, 33: 10-21.
CrossRef - Aubert, M., M. Hedde, T. Decaens, F. Bureau, P. Margerie and D. Alard, 2003. Effects of tree canopy composition on earthworms and other macro-invertebrates in beech forests of upper Normandy (France). Pedobiologia, 47: 904-912.
Direct Link - Deleporte, S., 2001. Changes in the earthworm community of an acidophilus lowland beech forest during a stand rotation. Soil Biol., 37: 1-7.
Direct Link - Gonzalez, G., T.R. Seastedt and Z. Donato, 2003. Earthworms, arthropods and plant litter decomposition in aspen and lodge pole pine forests in Colorado, USA. Pedobiologia, 47: 863-869.
Direct Link - Grant, C.D. and W.A. Loneragan, 2001. The effects of burning on the under story composition of rehabilitee bauxite mines bin Western Australia: Community changes vegetation succession. For. Ecol. Manage., 145: 255-277.
Direct Link - Hale, C.M., L.E. Frelich, P.B. Reich and J. Pastor, 2005. Effects of European earthworm invasion on soil characteristics in Northern Hardwood forests of Minnesota, USA. Ecosystem, 8: 911-927.
Direct Link - Haynes, R.J., C.S. Dominy and M.H. Graham, 2003. Effect of agricultural land use on soil organic matter status and the composition of earthworm communities in Kwazulu-natal, South Africa. Agric. Ecosyst. Environ., 95: 453-464.
Direct Link - Hedman, C.W., S.L. Grace and S.E. Ling, 2000. Vegetation composition and structure of Southern coastal plain pine forests: An ecological comparison. For. Ecol. Manage., 134: 233-247.
Direct Link - Hendrix, P.F. and P.G. Bonlen, 2002. Exotic Earthworm invasions in North America: Ecological and policy implications. Bioscience, 9: 1-11.
Direct Link - Huhtaq, V., 1979. Effects of liming and deciduous litter on earthworm populations of a spruce forest with an inoculation experiment on Allolobophora caliginous. Pedobiologia, 19: 340-345.
Direct Link - Jimens, J., P. Lavelle and T. Decaens, 2006. The efficiency of soil hand-sorting in assessing the abundance and biomass of earthworm community's. Its usefulness in population dynamics and cohort analysis studies. Eur. J. Soil Biol., 42: S225-S230.
Direct Link - Johnson-Maynard, J.L., R.C. Graham, L. Wu and P.J. Shouse, 2002. Modification of soil structural and hydraulic properties after 50 years of imposed chaparral and pine vegetation. Geoderma, 110: 227-240.
Direct Link - Kimmins, J.P., 1987. Forest Ecology: A Foundation for Sustainable Management. 1st Edn., The University of British Columbia, Columbia, pp: 596.
Direct Link - Neirynck, J.S., G. Mirtcheva and N. Lust, 2000. Impact of Tilia patyphyllos scop. Fraxinus excelsior L., Acer pseudoplatanusl. Quercus robur L. and Fagus sylvatica L. on earthworm biomass and physico-chemical properties of loamy topsoil. For. Ecol. Manage., 133: 275-286.
Direct Link - Parkinson, D. and S.E. Welke, 2003. Effects of Aporrectodea activity on seedling growth on Pseudotsuga menzeissi, nutrient dynamics and microbial activity in different forest soils. For. Ecol. Manage., 173: 169-186.
Direct Link - Ratty, M., 2004. Growth of Lumbericus terristris and Aporrectodea caliginosa in an acid forest soil and their effects on enchytraeid populations and soil properties. Pedobiologia, 48: 321-328.
Direct Link - Ratty, M. and V. Hutha, 2004. Earthworm communities in birch stand with different origin in central Finland. Pedibiologia, 48: 283-291.
Direct Link - Sautter, K.D., C.G. Brown, S.W. James, D.H. Pasini, N. Nunesand and P. Benito, 2006. Present knowledge on earthworm biodiversity in the state of Parana. Brazil. Eur. J. Soil Biol., 42: S296-S300.
Direct Link - Scheu, S. and M. Schaefer, 1998. Bottom-up control of the soil macro fauna community in a beech wood on limestone: Manipulation of food resources. Ecology, 79: 1573-1585.
Direct Link - Scheu, S., N. Schmitt, A.V. Tiunov, J.E. Newington and T.H. Jones, 2002. Effects of the presence and community composition of earthworms on microbial functioning. Oecologia, 133: 254-260.
Direct Link - Sheehan, L., J. Kirwan, L. Connolly and T. Bolger, 2006. The effects of earthworm functional group diversity on nitrogen dynamics in soils. Soil Biol. Biochem., 38: 2629-2636.
Direct Link