
A. Aynehband
Department of Agronomy and Plant Breeding, Faculty ofAgriculture,
Shahid Chamran University of Ahvaz, Ahvaz, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 80-85







ABSTRACT
A 2 year field study was carried out at Agricultural Faculty of Shahid Chamran University of Ahvaz (Iran), in order to evaluate the response of Amaranth cultivars and weed dynamic to N splitting methods. Three Amaranth cultivars (i.e., Amont, Trigin and Plainsman) were grown in three N splitting methods (i.e., Commonly, Equal and Semi-equal) that being applied at planting, 12th leaf appearance and stem elongation. A split plot design replicated three times was used which Amaranth cultivars and N splitting methods were arranged in main and sub plots, respectively. Results showed that the highest forage yield was obtained for Trigin with Equal N splitting (i.e., 31.2 t ha-1) and Plainsman with Semi-equal N distribution (i.e., 3.8 t ha-1) the lowest. Also, the maximum and minimum protein content (%) were obtained for Trigin with Equal N splitting (16%) and Amont with commonly N splitting (11.9%), respectively. It was found that just Trigin with Equal N splitting treatment was the best treatment for both forage quality and quantity yield. Moreover, the weed communities and dominant species changed in response to various N splitting methods and Amaranth cultivars traits. Plainsman with Semi-equal N splitting treatment was the unfavorable treatment for both crop yield and weed infestation. Based on these results it is recommend that N splitting method be applied mainly as an Equal form in Trigin amaranth cultivar, to enhance crop forage yield and reduce weed infestation.
PDF Abstract XML References Citation
How to cite this article
A. Aynehband, 2008. Cultivar and Nitrogen Splitting Effects on Amaranth Forage Yield and Weed Community. Pakistan Journal of Biological Sciences, 11: 80-85.
DOI: 10.3923/pjbs.2008.80.85
URL: https://scialert.net/abstract/?doi=pjbs.2008.80.85
DOI: 10.3923/pjbs.2008.80.85
URL: https://scialert.net/abstract/?doi=pjbs.2008.80.85
INTRODUCTION
Amaranth (Amaranthus sp.) as a new crop is an ancient pseudo-cereal originating in the Americas and can be used as a high-protein grain and forage crop or a leafy vegetable. Amaranth is a dicot, broadleaf, with C-4 photosynthesis and summer annual crop responding well to high sunlight and warm temperature. The plant height ranges from 91 to 274 cm and soil temperature above 15°C is required to establish a good plant stand. The two species of amaranth commonly grown are A. cruentus L. (2n = 34) and A. hypochondriacus L. (2n = 32) that can be used as a dual-purpose crop. The potential of amaranths as forage has not been fully studied but highly prized as forage crops because of their rapid growth rate, high protein content (the leaves, stem and head have a 15-24% protein) and good forage yield (Myers, 1996). Several studies have shown that yield differences of amaranth cultivars were due to nitrogen availability (Myers, 1998; Pospisil et al., 2006; Schulte et al., 2005) and individual cultivar traits (Henderson et al., 2000; Sleugh et al., 2001; Stordahl et al., 1999). To grow this alternative cereal efficiently, it is necessary to know the effect of nitrogen fertilization on its yield because nitrogen was found to be the primary limiting factor of amaranth production (Pospisil et al., 2006). Varieties of different amaranth species respond differently to the nitrogen amount applied. The author recorded low efficiency of nitrogen use by amaranth with the increase of soil nitrogen content. Therefore, higher nitrogen rate may have an adverse effect on the harvest due to increased plant height, lodging and protracted seed ripening (Elbehri et al., 1993). Also, some amaranth can have toxic levels of nitrate and oxalates that can result from nitrates in forages, especially if drought conditions accrue during a period of heavy nitrate uptake by plant (Sleugh et al., 2001). Similarly it is reported that although amaranth has some potential for use as forage, it is important to consider the potential for accumulation of excessive NO3 in the leaves due to high soil N. Leaf NO3 and certain other leaf properties increased in response to fertilizer rate of 50 and 100 kg N ha-1. But, the leaf NO3 levels fell rapidly as the season progressed (Myers, 1998). Generally, nitrogen uptake seems to be very efficient in amaranth. Without N fertilization, still 140 kg N ha-1 were taken up; more than double the amount present in the soil at sowing. This clearly exceeds comparable values for wheat and barley (Schulte et al., 2005). Differences in N use have been reported as function of genotype, N fertilizer timing and other factors, suggesting considerable opportunities for improving NUE by managing cropping system components (Lopez-Bellido et al., 2006). Report that the time of fertilizer N applications has a significant effect on the uptake of fertilizer N by the crop and the resulting partitioning of added N between soil and plant (Limaux et al., 1999). Moreover, the efficiency of N fertilizer use when applied as a topdressing in wheat is influenced by timing and fertilizer rate. Perhaps even more important than the optimum rate to use is the timing and splitting of nitrogen application. Splitting of N fertilizer application has been suggested as a strategy to improve crop N use efficiency (Lopez-Bellido et al., 2005).
From another point of view, for many producers, weed control is their biggest problem, because herbicides are not presently available for weed control in amaranth, weed control is accomplished with change in agronomic practices. For example, manipulation of crop fertilization is a promising cultural practice to reduce weed interference in crop (Angonin et al., 1996). Although nutrients clearly promote crop growth, in some cases, fertilizers benefit weeds more than crops. The increase in weed competition at higher N rate has been suggested to be related to an increase in the efficiency of nutrient accumulation and use by weed (Di Tomaso, 1995). Moreover, N application may influence the composition of the weed flora, suggesting that the effect of N fertilizer is not direct but depends on the density and competitiveness of the crop (Scursoni and Arnold, 2002).
There is only limited information about amaranth agronomic characters in Asia. However, sensitivity of Amaranth cultivars to high N availability, absent of specific herbicide and different response of amaranth and weed to N fertilizer are some of the most important agronomic problems for this crop. Therefore, the objective of this study was to evaluate of how N splitting methods influence on Amaranth cultivars yield, as well as its effects on weed community.
MATERIALS AND METHODS
This field experiment was carried out during the two successive summer seasons of 2005 and 2006 at the experimental farm of Agricultural Faculty of Shahid Chamran University, Ahvaz, Iran (31°N, 48°E, with elevation at 20 m). The soil texture of the experimental site was clay loam. Mean air temperature from June to September was 40°C, with no precipitation in this period. The experimental design was a randomized complete block with a split plot arrangement with three replications and nine plots per replication. The main plots were devoted to amaranth cultivars while the sub-plots were devoted to N splitting. The area of the sub plot was 13.5 m2 consisting of six rows 75 cm width and 3 m length. Plant population was 130000 plants ha-1. The amaranth cultivars used were A. cruentus cv. A mont and Trigin-G6 and A. hypochondriacus cv. Plainsman that were planted and harvested on 23 July and 2 October, respectively. N splitting was applied in various proportions among planting, 12th leaf appearance and stem elongation including: 25-0-75% (Common); 33-33-33% (Equal) and 25-50-25% (Semi-equal) distribution. Nitrogen fertilizer was applied at rate of 110 kg N ha-1. The normal agricultural practices for growing amaranth were followed as recommended by Thomas Jefferson Agricultural Institute, Columbia, MO. Plant harvest carried out manually at the flowering stage. Samples were dried before weighing. Above ground biomass yield, leaf and stem weight, protein content (%) and plant height were determined for each plot. Weeds were sampled at two weeks after each N splitting treatment. All the weeds in 1 m2 quadrate per plot were removed by hand, sorted by species, counted, dried and weighed. Total weed density and biomass (i.e., weed m2), total density and biomass of broadleaf and annual grass weed species and weed dominant species were determined. Data were processed by the analysis of variance the MSTAT-C program and treatment means were separated with Duncan test at p<0.05 (average means of two years data was used in tables).
RESULTS AND DISCUSSION
Forage yield: All forage yield parameters significantly changed depending on cultivars and N splitting application (Table 1). Trigin and plainsman cultivars with commonly and semi-equal N splitting had the highest and lowest plant height, respectively. In addition to the effect of amaranth cultivars, the plant height value had the lowest response to the two-part N splitting than the three parts N splitting. Trigin had the greatest plant height in all N splitting methods. In contrast, plainsman was less sensitive to N splitting for this factor. The stem and leaf weight were found highest for Trigin with equal N splitting than other treatments. It`s indicated that more than genotype effect (i.e., amaranth cultivars), the equal distribution of N topdressing had a clear influence on better nutrient absorption and vegetative growth in amaranth. Response of Plainsman to N splitting was very lower than Amont and Trigin cultivars that were similar behavior to the plant height for those cultivars. Elbehri et al. (1993) indicated that plant height proved to be very responsive to N fertilization. Similarly, Myers (1998) cited Amaranthus cruentus L. was the tallest cultivar, followed by Plainsman and its plant height was particularly responsible to N. Also, in many cases, stem weight was higher than leaf weight, the exception being Amont and Plainsman with semi-equal N distribution. Some researcher had found that higher dry matter content
| Table 1: | Forage yield and yield components of amaranth cultivars as affected by N splitting methods |
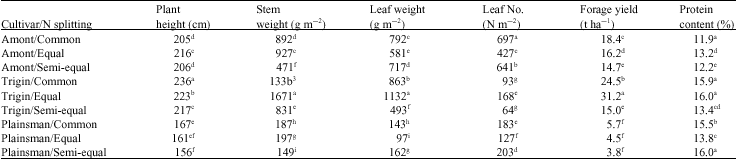 | |
| Means with the same letter(s) within the column are not significantly different at 5% probability level | |
in the flowering stage (especially higher stem weight) indicates the ability of plants to produce higher total dry matter (Pospisil et al., 2006). However the data here indicated that the highest and lowest leaf weight and leaf number were not obtained from similar treatments (Table 1), so, leaf thick change in response to these differences (data not shown). These data underlines the need to take into account interaction between genotype x agronomic methods and showed that some agronomic methods (i.e., N splitting) have different effects on crop genotype (i.e., amaranth cultivars) and vice versa.
Forage yield of amaranth cultivars showed a different response to N splitting (Table 1). The highest yield quantity (i.e., 31 t ha-1) was obtained by Trigin with equal distribution of N topdressing, but it`s reduced by change in N splitting methods (i.e., 32 to 15 t ha-1). So, Trigin cultivar was very sensitive to N splitting pattern. In contrast, forage yield of Plainsman was unaffected by N splitting methods (i.e., the lowest among the three cultivars). In the present experiment, Trigin with the equal N splitting had the greatest forage yield. This periphery can be attributed to both higher stem weight (i.e., 1671 g m-2) and leaf weight (i.e., 1132 g m-2) compared to other yield parameters. These results are in general agreement with other studies that emphasize on the effect of genotype and agronomic practices on crop yield (Henderson et al., 2000; Myers, 1996; Stordahl et al., 1999), but also studies amaranth in tropical regions have shown inconsistent yield responses to N availability, reflecting the diversity of environments and conditions under which the studies have been conducted (Myers, 1998).
Although, Trigin with equal N splitting had the highest protein content (similar to its forage yield), Plainsman with semi-equal N distribution also had protein content (%) just as much as Trigin with equal N splitting, in spite of its lowest forage yield. It seemed that leaf weight was more effective than leaf number in protein content (%). However, it is reported that protein content (%) of amaranth cultivars was affected by both leaf weight and number (Sleugh et al., 2001). In fact, Trigin with equal N splitting was the best treatment for yield quantity; Trigin with both common and equal along with Plainsman with semi-equal N splitting were the best treatments for yield quality; but it was just Trigin with equal N splitting that proved significantly superior for both yield quality and quantity. Some of these differences were due to individual genotype effects and others associated with N splitting methods. In other words, with managing of N application in time, forage quality could be improved not only in high yielding cultivar (i.e., Trigin) but also in lower yielding cultivar (i.e., Plainsman). Similarly, it is reported that the forage nutritive value of amaranth is equal to or better than commonly used forages, but quality parameters of amaranth cultivars (i.e., protein content and dry matter digestibility) were different (Pospisil et al., 2006; Schulte et al., 2005). Walters et al. (1988) showed that forage quality of several amaranth accessions declined linearly with changing in nitrogen availability levels. This suggested that nitrogen management could be an option in improving the forage quality of amaranth. The positive effect of splitting N fertilizer was also reported by Limaux et al. (1999) and Lopez-Bellido et al. (2006).
Weed dynamic: Amaranth cultivars and N splitting had a significant effect on weed community (Table 2). Maximum broadleaf weed density was obtained in Plainsman with semi-equal N splitting. Also, semi-equal N distribution method provided the highest broadleaf weed biomass in all amaranth cultivars. For each cultivar, the greatest broadleaf weed biomass was achieved in the highest broadleaf weed density treatment. Total weed density and biomass were greatest in Plainsman with semi-equal N treatment that may be related to agronomic traits of individual amaranth cultivar. Because this cultivar (i.e., Plainsman) had the lowest plant height, leaf number, leaf weight and also its canopy was closed later than the others. Therefore, poor establishment of Plainsman associated with semi-equal distribution of N, thus probably contributed most to increase in emergence of total weed species at this treatment. It appears that the
| Table 2: | Effect of N splitting methods on the weed density and biomass in amaranth cultivars |
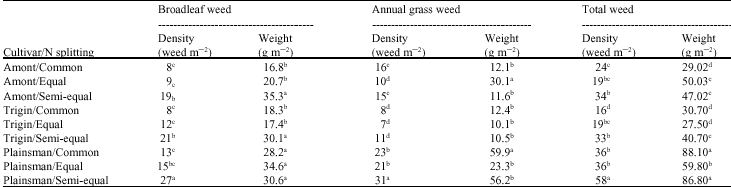 | |
| Means with the same letter(s) within the column are not significantly different at 5% probability level | |
earlier N is available, the more beneficial it is for crop`s competitive ability. In contrast, if N is available at late crop stage, the weed`s growth is favored (Scursoni and Arnold, 2002). This situation will exceed for crop like amaranth which grows slowly during the first few weeks and plant growth is not vigorous early in the season (Myers, 1998). Similarly, split application of N fertilizer may be a useful practice for managing weed in sweet corn. Early-season soil N levels are kept intentionally lower in a split application management system because sweet corn demand for N at this time is low, whereas the potential for loss of excess N from the system is high (Davis and Liebman, 2001). In another study, it was reported that the split application of nitrogen caused slightly higher increases of sterile oat dry weight and total nitrogen and greater grain yield reduction of wheat grown with oat, compared to that of a single application (Dhima and Eleftherohorinos, 2001).
Weed dominant species: More scrutiny evaluation of weed communities showed that cultural approaches (amaranth cultivars and N splitting) also had a significant effect on diversity of dominant weed species. In most N splitting methods, Plainsman had the greatest density and frequency of broadleaf weed dominate species (Table 3). Also, this amaranth cultivar with semi-equal N treatment had a higher density and biomass of weed dominate species than other treatments. Evidently, the presence of unfavorable amaranth cultivar (Plainsman), in spite of N splitting methods, resulted in just one weed species (Chenopodium album L.) became dominant in broadleaf weed community with the same phenological stage (i.e., in the budding stage). Conversely, in more favorable amaranth cultivars (i.e., Amont and Trigin), agronomic traits with a concomitant N splitting methods resulted in diversification of dominant broadleaf species depending on density, biomass and phonological stage. Therefore, it`s remarkable that the influence of N splitting methods on dominant broadleaf community was higher in good amaranth cultivar stands in comparison with the poor ones. These results are in general agreement with other studies indicating that changes in weed community composition are the result of selection pressures imposed by agronomic practices (Derksen et al., 2002). For example, the ability of a species to better utilize available nutrients can also provide an advantage in competition for water and light. Therefore, its can be dominant (Di Tomaso, 1995). Also, it is reported that crop cultivars vary in a number of developmental characteristics, including stature, canopy development and leaf parameters. These qualities can have a dramatic effect on competitiveness in the presence of weed species (Angonin et al., 1996; Di Tomaso, 1995). Other research has shown that the level of N availability, through its effect on weed growth and the resultant seed rain, potentially could influence the species composition of weed community. Therefore, agronomic practices that influence the competitive ability of a crop also can select against less competitive weed species (Stevenson et al., 1997).
Dominant annual grass weed species was also affected by amaranth cultivars and N splitting treatments (Table 4). Under such conditions, the highest density and biomass of weed dominant species with frequency of 70.9 and 84.62% were obtained in Plainsman cultivar with semi-equal N treatment, respectively. Data showed that annual grass dominant species was more affected by amaranth cultivars than N splitting methods, because each amaranth cultivar had a specific weed dominant species (Sorghum halepense L. in Amont, Cyperus rotundus L. Trigin and Echinochloa crus-galli (L.) Beauv. in Plainsman). Although each amaranth cultivar had an individual dominant weed species, their phenological stages were different. These kinds of responses could be attributed to the effect of N splitting method on pattern of N availability to weeds. In other words, varying in N availability led to different weed phenology. These results are in the same line of the finding reported that weed communities are constantly evolving in response to crop management practices. Fertilization alters soil fertility,
| Table 3: | Dominant broadleaf weed species parameters in amaranth cultivars as affected by N splitting methods |
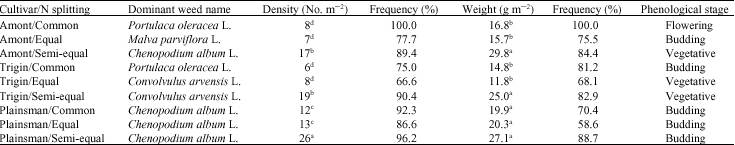 | |
| Means with the same letter(s) within the column are not significantly different at 5% probability level | |
| Table 4: | Dominant annual grass weed species parameters in amaranth cultivars as affected by N splitting methods |
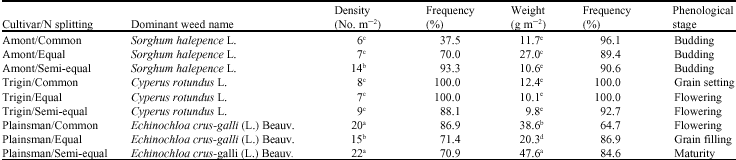 | |
| Means with the same letter(s) within the column are not significantly different at 5% probability level | |
which affects not only crop growth but also composition and growth of associated weeds. Therefore, understanding the shifts in weed community composition under different fertilization availability would help in designing effective crop management program (Yin et al., 2005). Other studies suggested that since seed production is correlated with individual biomass, the species adapted to a form of N availability would produce more offspring than other weeds and increase in abundance in the weed community (Angonin et al., 1996).
CONCLUSION
The results of this research indicated that amaranth with production of high quality and quantity of forage yield can be established as a prominent and profitable alternative forage crop in our cropping systems. Amaranth cultivars with different agronomic traits were affected by N splitting methods. Trigin cultivar and Equal N splitting produced highest forage yield (i.e., 31.2 t ha-1) and protein content (i.e., 16%), respectively. Also, it was observed that weed flora was changed depending on amaranth cultivars and N splitting methods. Commonly and Semi-equal N distribution, were more favorable for weeds than crops growth, respectively. In addition, weed phenology was very responsive to N splitting methods. These findings indicate that some common agronomic practices although obviously useful for crop yield will also stimulate weed infestation, too. Therefore, crop production efficiencies can be gained through appropriate fertilizer timing methods and support this concept that producers could enhance crop yield and concomitantly reduce weed problem by choosing more conscious agronomic practices.
REFERENCES
- Angonin, C., J. Caussanel and J. Meynard, 1996. Competition between winter wheat and Veronica hederifolia: Influence of weed density and the amount and timing of nitrogen application. Weed Res., 36: 175-187.
Direct Link - Davis, A.S. and M. Liebman, 2001. Nitrogen source influences wild mustard growth and competitive effect on sweet corn. Weed Sci., 49: 558-566.
CrossRefDirect Link - Derksen, D., R. Andaeson, R. Blackshow and B. Maxwell, 2002. Weed dynamic and management strategies for cropping system in the Northern Great Plains. Agron. J., 94: 174-185.
Direct Link - Dhima, K.V. and I.G. Eleftherohorinos, 2001. Influence of nitrogen on competition between winter cereals and sterile oat. Weed Sci., 49: 77-82.
CrossRefDirect Link - Di Tomaso, J., 1995. Approaches for improving crop competitiveness through the manipulation of fertilization strategies. Weed Sci., 43: 491-497.
Direct Link - Elbehri, A., D. Putnam and M. Schmitt, 1993. Nitrogen fertilizer and cultivar effects on yield and nitrogen use efficiency of grain amaranth. Agron. J., 85: 120-126.
Direct Link - Henderson, T., B. Johnson and A. Schneiter, 2000. Row spacing, plant population and cultivar effects on grain amaranth in the Northern Great Plains. Agron. J., 92: 329-336.
Direct Link - Limaux, F., S. Recous, J. Meynard and A. Guchert, 1999. Relationship between rate of crop growth at date of fertilizer N application and fate of fertilizer N applied to winter wheat. Plant Soil, 214: 49-59.
CrossRef - Lopez-Bellido, L., R.J. Lopez-Bellido and R. Redondo, 2005. Nitrogen efficiency in wheat under rainfed Mediterranean conditions as affected by split nitrogen application. Field Crop Res., 94: 86-97.
CrossRefDirect Link - Lopez-Bellido, L., R.J. Lopez-Bellide and F.S. Lopez-Bellido, 2006. Fertilizer nitrogen efficiency in durum wheat under rainfed Mediterranean conditions: Effect of split application. Agron. J., 98: 55-62.
Direct Link - Pospisil, A., M. pospisil, B. Varga and Z. Svecnjak, 2006. Grain yield and protein concentration of two amaranth species (Amaranthus spp.) as influenced by the nitrogen fertilization. Eur. J. Agron., 25: 250-253.
CrossRefDirect Link - Erley, G.S.A., H.P. Kaul, M. Kruse and W. Aufhammer, 2005. Yield and nitrogen utilization efficiency of the pseudocereals amaranth, quinoa and buckwheat under differing nitrogen fertilization. Eur. J. Agron., 22: 95-100.
CrossRefDirect Link - Scursoni, J. and R. Arnold, 2002. Effect of nitrogen fertilization timing on the demographic processes of wild oat in barley. Weed Sci., 50: 616-621.
Direct Link - Sleugh, B., K. Moore, E. Brummer, A. Knapp, J. Russell and L. Gibson, 2001. Forage nutritive value of various amaranth species at different harvest dates. Crop Sci., 41: 466-472.
Direct Link - Stevenson, F., A. Legere, R. Simard, D. Angers, D. Pageau and J. Lafond, 1997. Weed species diversity in spring barley varies with crop rotation and tillage, but not with nutrient source. Weed Sci., 45: 798-806.
Direct Link - Stordahl, J., C. Sheaffer and A. DiCostanzo, 1999. Variety and maturity affect amaranth forage yield and quality. J. Prod. Agric., 12: 249-253.
Direct Link - Walters, R.D., D.L. Coffey and C.E. Sams, 1988. Fiber, nitrate and protein content of Amaranthus accessions as affected by soil nitrogen application and harvest date. Hortscience, 23: 338-341.
Direct Link - Yin, L., Z. Cai and W. Zhong, 2005. Changes in weed composition of winter wheat crops due to long-term fertilization. Agric. Ecosyst. Environ., 107: 181-186.
CrossRef