
Yesim Koc
Egitim FakUltesi, Sinop Uni2versi2tesi2, 57000 Korucuk/Sinop, TUrkiye
Adem GUlel
Fen Edebiyat FakUltesi, Ondokuz Mayi2s Uni2versi2tesi2, Kurupelit/Samsun, TUrkiye
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 5 | Page No.: 733-739







ABSTRACT
Variations in protein and carbohydrate levels correlated with the age and sex of Galleria mellonella (Linnaeus, 1758) (Lepidoptera: Pyralidae) in constant lightness and darkness were investigated. Tests were conducted under laboratory conditions at 28 ±2 °C temperature and 65 ± 5% relative humidity. Insects were fed on combs without honey. Protein level in 100 mg of adults increased in the first days of adult life of females in connection with their age and then decreased. No difference was observed in males. Carbohydrate level in 100 mg of adults increased in both sexes in connection with their age. In all tests carbohydrate and protein levels of females were found higher than males. Protein and carbohydrate levels of adult G. mellonella varied in connection with the photoperiod regimes implemented. Decrease in the nutrient levels was observed in constant darkness.
PDF Abstract XML References Citation
How to cite this article
Yesim Koc and Adem GUlel, 2008. Age and Sex Related Variations in Protein and Carbohydrate Levels
of Galleria mellonella (Linnaeus, 1758) (Lepidoptera: Pyralidae)
in Constant Lightness and Darkness. Pakistan Journal of Biological Sciences, 11: 733-739.
DOI: 10.3923/pjbs.2008.733.739
URL: https://scialert.net/abstract/?doi=pjbs.2008.733.739
DOI: 10.3923/pjbs.2008.733.739
URL: https://scialert.net/abstract/?doi=pjbs.2008.733.739
INTRODUCTION
Protein, lipid and carbohydrate metabolism plays role in many vital activities of insects. Many other factors such as sex (AktUmsek, 1996; Ito and Nakata, 1998), age (Jacome et al., 1995; Seker and Yanikoglu, 1999; Akman, 2004), developmental stages (Bozkurt, 2003), diapause (Pullin, 1992), nutrient quality and level (Yanikoglu, 1985; Jacome et al., 1995; Ozalp and Emre, 1998; Socha et al., 1998; George et al., 2002), seasonal conditions (Ito, 1989; Ito and Nakata, 1998), temperature (Varer, 2005), host type in some species (AktUmsek, 1996; Akman, 2004), sexual activity (Warburg and Yuval, 1996), use of insecticide (Sak, 2004), photoperiod (Shuxia and Adams, 2000; Nakasuji and Mizumoto, 2001; Barsagade and Tembhare, 2002; El-Aw, 2003) are effective on the levels of these substances.
Adult insects need certain chemicals and energy in large quantities to maintain vital activities such as mating, food searching, oviposition and parasitism. Therefore, having basal nutrients such as carbohydrates, lipids and proteins in certain quantities is a necessity (England and Evans, 1997; Olson and Andow, 1998; Ozalp and Emre, 1998; Meats and Leighton, 2004). Necessary nutrients can be stored in larva or pupa stages or adults can synthesize them by taking related precursor compounds. Morales-Ramos et al. (1996) found that Catolaccus grandis (Hymenoptera: Petromolidae) females without a carbohydrate source die in 2.5 days at 30°C but when they are given a mixture of glucose and fructose in equal quantity each, they can survive 18.27 days under the same conditions. Many studies on Trichogramma show that carbohydrate source in adult nutrition prolong life (Olson and Andow, 1998).
Proteins are the last option among the substances to be used as fuel. Proteins are effective especially on metamorphosis, growth, cocoon and cuticula formation and flying. Protein level varies especially before or during metamorphosis in the developmental stage (Socha and Sula, 1992; Shuxia and Adams, 2000; El-Aw, 2003; Meats and Leighton, 2004).
Many insects need sugars for metabolic processes or for using them as the precursor compounds. These are sensitive to carbohydrate levels (England and Evans, 1997; Olson and Andow, 1998; Olson et al., 2000). Carbohydrate sources, which are used by insects in the nature, such as nectar can be instantly used in metabolic activities or converted to glycogen or trehalose and reserved for using later (Olson et al., 2000).
Some insects use carbohydrates as the primary source of energy and keep excess carbohydrate as lipid and protein. Generally Diptera and Hymenopera use carbohydrates as a main energy source (Bailey, 1975) Factors such as photoperiod (Pullin and Wolda, 1993), age (Seker and Yanikoglu, 1999), nutrition (Yanikoglu, 1985; Ozalp and Emre, 1998) and host (Akman, 2004) are effective on carbohydrate metabolism.
G. mellonella is a harmful species for combs and its larvae are known as causing serious damages in beekeeping (Chang and Hsieh, 1992; Haewoon et al., 1995; Charriere and Imdorf, 1999; Hood et al., 2003). There are not many studies available regarding the effects of photoperiod, age and sex on carbohydrate and protein levels of this insect. Since significant variations in carbohydrate and protein levels can be observed in connection with age and sex, the effects of constant lightness and darkness as well as the effects of age and sex on quantities of the substances in question will be focused in this study.
MATERIALS AND METHODS
Large wax moth Galleria mellonella (Linnaeus, 1758) (Lepidoptera: Pyralidae) was employed for the tests. Tests were conducted under laboratory conditions at 28±2°C; 65±5% relative humidity and in constant lightness or darkness. Studies started by establishing successive laboratory stock cultures of G. mellonella. Methods of Koc and GUlel (2006) were used for establishing the stock culture, distinguishing adults by sexes and grouping by age.
Insects were let to proliferate in jars at predefined constant temperature and humidity in constant darkness (DK) or constant lightness (DA). One day old adults were weighed and stocked on the first day of their adult life. Ten adults matured on the same day from five and fifteen days old groups put into a jar and were fed on comb without honey. Five of them were weighed five days later and another five of them 15 days later, respectively. Processes for the mentioned three age groups were repeated three times with samples taken from the population at different times. For each analysis, 15 adult insects from a certain age group were weighed and stocked in 1.5 mL Eppendorf tubes at -50°C until they had been analyzed.
Biochemical analysis: For carbohydrate and protein analysis, one, five and fifteen days old adults stocked at -50°C were used.
Protein analysis: First of all a standard protein graph was created for specifying protein values that will be obtained through protein analysis. About 0.1% bovine serum albumin was used. The standard stock solution in 1 mg mL–1 concentration was prepared. Then standard protein solutions in 10, 20, 25, 30, 40, 50, 75 and 100 μg mL–1 concentrations were prepared through serial dilutions of the standard stock solution.
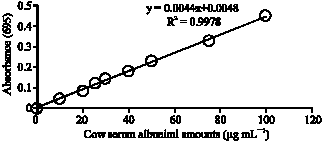 | |
| Fig. 1: | Standard protein graph |
Lowry method was applied to these solutions and the absorbance values monitored by a spectrophotometer at 695 nm wavelength against blank. These processes were repeated three times for each concentration of the standard solution. Standard protein graph was created by using the absorbance values obtained (Fig. 1).
Total protein analysis in the stocked samples was based on the method developed by Lowry et al. (1951). The samples weighed and kept at -50°C were transferred into larger tubes for homogenization after they had been kept at room temperature for a while. Each tube had one insect in it and 5 mL work buffer was added to the tubes. Each insect was homogenized at 8,000 rpm for 7 min. The homogenate was centrifuged at room temperature at 3,500 rpm for 15 min. About 100 μL of the supernatant generated in the tube at the end of the centrifuging was processed with Lowry method and monitored with a spectrophotometer at 695 nm wavelength against blank. The monitored absorbance values were evaluated by using the standard protein graph.
Carbohydrate analysis: First of all a standard carbohydrate graph was created for specifying carbohydrate values that will be obtained through carbohydrate analysis. 0.1% glucose solution was used. Then glucose solutions in 25, 50, 75 and 100 μg mL–1 concentrations were prepared through serial dilutions of the stock solution. These standard solutions were put into reaction with Anthron reactive agent and heated at 90°C for 15 min. The color was changed. Then the tubes were cooled, stirred and the absorbance values were monitored at 695 nm wavelength with a spectrophotometer. These processes were repeated three times (Fig. 2).
Van Handel (1985) Anthrone test was employed for measuring the carbohydrate level in the samples stocked for analysis. The samples stored in a deep freezer at -50°C were kept at room temperature for a while and each of them homogenized at 8000 rpm for 7 min in 2.5 mL 2% Sodium Sulfate. One hundred and fifty microliter supernatant was taken from the samples centrifuged at room temperature at 16000 rpm for 2 min after they had been stirred.
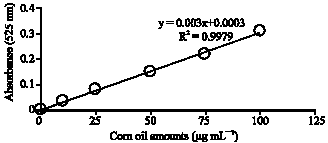 | |
| Fig. 2: | Standard carbohydrate graph |
2 mL Anthrone was added to the samples and the tubes heated at 90°C for 15 min. The color was changed. Then the tubes were cooled on ice, stirred and monitored at 625 nm against blank. Carbohydrate levels were measured by using the standard carbohydrate graph.
Statistical assessment of the data collected: One Way Analysis of Variance (ANOVA) was used for the comparison of more than two groups. Averages were assessed by using Student-Newman-Kuel (SNK) Test, when the test results were significant. Independent Two Samples t-test was employed for the comparison of two groups and a = 0.05 confidence limit was taken as basis.
RESULTS
Other results in connection with the protein analysis: Result of age and sex related protein level analysis on adult G. mellonella kept in constant darkness is given in Table 1.
SH-standard error: As it is shown in Table 1, protein levels are different in male and female adults. In females, the protein level was increased in the first five days of adult life but then decreased afterwards. In males, the protein level kept increasing after the fifth day of adult life. The average protein level was 4.31 mg in 100 mg of adult females one day older than adults from the same age group, while the males from the same age group had 4.01 mg protein, in average.
The average protein levels of five days old females and males were 5.51 and 5.11 mg, respectively, while the levels were 4.91 and 5.43 mg in fifteen days old ones. The difference of protein levels between sexes for one and five days old adults are statistically insignificant (p>0.05), whereas the difference between sexes is significant for fifteen days old adults (p<0.05). The difference in protein levels between the first day of adult life and the 15th day is significant (p<0.05) for both sexes of G. mellonella adults.
| Table 1: | Protein level of adult G. mellonella in constant darkness |
 | |
*Average of 3 repeated processes each with 5 beings, **Significance level between the protein levels of males and females from the same age group, Average differences in the same column with the same letter(s) are not significant (p<0.05) | |
| Table 2: | Protein level of G. mellonella in constant light |
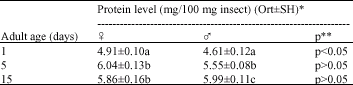 | |
*Average of 3 repeated processes each with 5 beings, **Significance level between the protein levels of males and females from the same age group, Average differences in the same column with the same letter(s) are not significant (p<0.05) | |
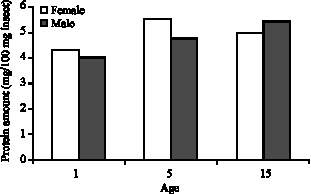 | |
| Fig. 3: | Age and sex related protein level of G. mellonella in constant darkness |
Results of age and sex related protein level analysis on adult G. mellonella kept in constant lightness are given in Table 2.
In females, the protein level was increased in the first five days of adult life but then decreased afterwards (Table 2). In males, the protein level kept increasing after the fifth day of adult life. The average protein level was 4.91 mg in 100 mg of adult females one day older than adults from the same age group, while the males from the same age group had 4.61 mg protein, in average. The difference of protein levels between sexes for five and fifteen days old adults are statistically insignificant (p>0.05), whereas the difference between sexes is significant for one days old adults (p<0.05). The difference in protein levels the first day of adult life and the 15th day is significant (p<0.05) for both sexes of G. mellonella adults.
| Table 3: | Carbohydrate level of adult G. mellonella in constant darkness |
 | |
*Average of 3 repeated processes each with 5 beings, **Significance level between the protein levels of males and females from the same age group, Average differences in the same column with the same letter(s) are not significant (p<0.05) | |
| Table 4: | Carbohydrate level of adult G. mellonella in constant light |
 | |
*Average of 3 repeated processes each with 5 beings, **Significance level between the protein levels of males and females from the same age group, Average differences in the same column with the same letters are not significant (p<0.05) | |
| |
| Fig. 4: | Age and sex related protein level of G. mellonella in constant light |
Results in connection with the carbohydrate analysis: Results of age and sex related carbohydrate level analysis on adults G. mellonella are given in Table 3 and 4.
SH-standard error: As it is seen in Table 3 carbohydrate levels in G. mellonella adults increased in both sexes connection with their ages. The difference of carbohydrate levels between in both sexes and in all age groups is statistically significant (p<0.05).
The carbohydrate levels in G. mellonella adults increased in both sexes connection with age (Fig. 4).
The carbohydrate levels in G. mellonella adults increased in both sexes connection with age (Table 4). The difference of carbohydrate levels between in both sexes and in all age groups is statistically significant (p<0.05).
DISCUSSION
Age and protein level relations, variations in protein level accompanied by aging and different protein necessities of females and males were studied by various researches (Shuxia and Adams, 2000; Akman, 2004; Meats and Leighton, 2004). In this study, variations in protein levels of adults in accordance with the adult age are observed (Table 1, Fig. 3). For this variation it was observed that in females increase in the beginning was followed by a decrease, whereas the increase kept going in males. Protein levels of females first increased in the beginning of adult life for high rate of egg production. However, as the reproduction potential declined by aging in the following days of life, the protein demand necessary for the egg production was also declined. Protein levels of adults were decreased when this condition was united with the decreased anabolic reactions. Since the protein used by males for sperm production is not consumed in large quantities as by females, the decrease observed in the protein level of males was less than the decrease observed for females. The results obtained from this study of the age related protein level variation are in agreement with the results of other researches. In a research with Spodoptera littoralis (Lepidoptera: Noctuidae) the effects of host-plant, photoperiod, day time, developmental stage and sex on the protein band number and concentration were studied and it was found that the protein band number of the last instar larvae change (El-Aw, 2003).
Age-related changes in hemolymph free amino acids and proteins were examined by Shuxia and Adams (2000), when Colorado Potato Beetle-Leptinotarsa decemlineata was reared under both short-day (8:16) (L:D) and long-day (17:7) (L;D) conditions. Under a short-day photoperiod, the total free amino acid concentration in the hemolymph increased gradually up to 20 days of adult life, but the long-day beetles showed marked increases during the first 10 days and then decreased afterwards. Proline, glutamine and valine were the most abundant free amino acids in both sexes.
In a study with Bactrocera tryoni (Diptera: Tephritidae) it was revealed that for egg production, protein consumption of sexually matured adults are higher than non-sexually matured females (Meats and Leighton, 2004). High protein levels of the females began to decrease especially after the second week. The increase in the protein level until becoming sexually matured and then the decrease in the protein level with the start of egg production and age related protein issues, which were observed in this study with G. mellonella, are in line with other studies with different insects. Protein levels derived from adults were found different in males and females as well as in lightness and darkness.
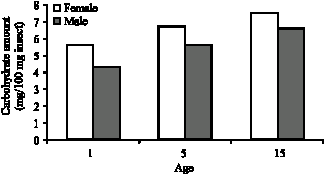 | |
| Fig. 5: | Age and sex related carbohydrate level in G. mellonella in constant darkness |
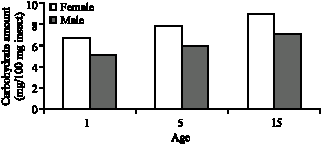 | |
| Fig. 6: | Age and sex related carbohydrate level of G. mellonella in constant light |
These differences might be due to photoperiod induced endocrine imbalance changing the protein level.
In this study with G. mellonella the relation between adult age and carbohydrate levels and age related gradual increase in carbohydrate level were observed (Table 2, Fig. 4). This increase might be in connection with the food as well as the inhibition of extreme carbohydrate consumption in the body due to Lepidoptera`s lipid preference as the primary energy source. Increase in the carbohydrate level is not related to the glucose synthesis from lipids because animals do not have the glyoxylate converting enzyme and glucose can not be synthesized from lipids. Besides, age related carbohydrate increase might be in connection with the components of the wax, which is the food, as they are employed in glucose synthesis. Carbohydrate level of females, which is higher than males, can be linked to females` ability of accumulating and synthesizing more carbohydrate from the food they were given and higher carbohydrate consumption of males. It is proven by various researchers (Olson et al., 2000; Giron and Casas, 2003) that in pre-adult and post-adult phases, females generally keep more protein, carbohydrate and lipids than males. Age related increase in glycogen levels were observed in the studies on age related carbohydrate metabolism with other insects (Seker and Yanikoglu, 1999; Akman, 2004; Varer, 2005).
Variations in nutrient levels between G. mellonella adults kept in constant lightness and constant darkness depend on the variations between insects` activity and energy requirement according to day length (Fig. 5-8).
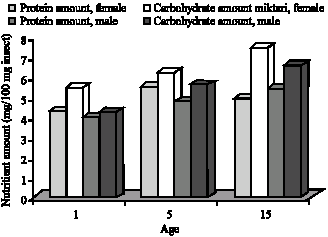 | |
| Fig. 7: | Age and sex related carbohydrate and protein levels of G. mellonella in constant darkness |
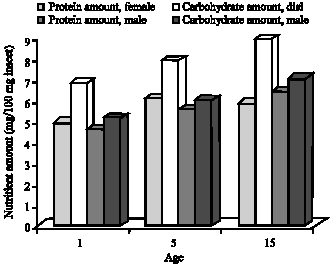 | |
| Fig. 8: | Age and sex related carbohydrate and protein levels of G. mellonella in constant light |
In this study, age and sex related variations in the insect metabolism in two different photoperiods and the effects of these variations on protein and carbohydrate levels were observed. The results of our study are in line with the results of other studies on other insects in this scope (Seker and Yanikoglu, 1999; Shuxia and Adams, 2000; Olson et al., 2000; El-Aw, 2003; Giron and Casas, 2003; Akman, 2004; Meats and Leighton, 2004; Varer, 2005).
REFERENCES
- Bozkurt, K., 2003. Phospholipid and triacylglycerol fatty acid compositions from various development stages of Melanogryluss desertus Pall. (Orthoptera: Gryliidae). Türk. J. Biol., 27: 73-78.
Direct Link - Giron, D. and J. Casas, 2003. Lipogenesis in adult parasitic wasp. J. Insect. Physiol., 49: 141-147.
CrossRef - Hood, W., P.M. Horton and J.M. McCreadie, 2003. Field evaluation of the imported fire ant (Hymenoptera: Formicidae) for the control of wax moths (Lepidoptera. Pyralidae) in stored honey bee comb. J. Agric. Urban. Entomol., 20: 93-103.
Direct Link - Koc, Y. and A. Gulel, 2006. Effects of different photoperiod regimes on total lipid amounts of Galleria mellonella (Linnaeus, 1758)(Lepidoptera: Pyralidae) adults at different age. Turk. Entomol., 30: 209-222.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Meats, A. and S.M. Leighton, 2004. Protein consumption by mated, unmated, sterile and fertile adults of the Queensland fruit fly, Bactrocera tryoni and its relation to egg production. Physiol. Entomol., 29: 176-182.
CrossRefDirect Link - Olson, D.M., H. Fadamiro, J.G. Lundgren and G.E. Heimpel, 2000. Effects of sugar feeding on carbohydrate and lipid metabolism in a parasitic wasp. Physiol. Entomol., 25: 17-26.
CrossRef - Shuxia, Y.Y. and T.S. Adams, 2000. Effect of pyriproxyfen and photoperiod on free amino acid concentrations and proteins in the hemolymph of colorado potato beetle, Leptinotarsa decemlineata. J. Insect. Physiol., 46: 1341-1353.
Direct Link - Van Handel, E., 1985. Rapid determination of total lipids in mosquitoes. J. Am. Mosquito Control Assoc., 1: 302-304.
PubMed