
Yunus Shukor
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Burhanuddin Shamsuddin
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Othman Mohamad
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Khalid Ithnin
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Nor Aripin Shamaan
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Mohd. Arif Syed
Department of Biochemistry, Faculty of Biotechnology and Biomolecular Sciences, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 4 | Page No.: 672-675







ABSTRACT
In this research, we modify a previously developed assay for the quantification molybdenum blue to determine whether inhibitors to molybdate reduction in bacteria inhibits cellular reduction or inhibit the chemical formation of one of the intermediate of molybdenum blue; phosphomolybdate. We manage to prove that inhibition of molybdate reduction by phosphate and arsenate is at the level of phosphomolybdate and not cellular. We also prove that mercury is a physiological inhibitor to molybdate reduction. We suggest the use of this method to assess the effect of inhibitors and activators to molybdate reduction in bacteria.
Key words: Molybdate reduction, E. cloacae strain 48, molybdenum blue, inhibitors
PDF Abstract XML References Citation
How to cite this article
Yunus Shukor, Burhanuddin Shamsuddin, Othman Mohamad, Khalid Ithnin, Nor Aripin Shamaan and Mohd. Arif Syed, 2008. A Method to Study the Effects of Chemical and Biological
Reduction of Molybdate to Molybdenum Blue in Bacteria. Pakistan Journal of Biological Sciences, 11: 672-675.
DOI: 10.3923/pjbs.2008.672.675
URL: https://scialert.net/abstract/?doi=pjbs.2008.672.675
DOI: 10.3923/pjbs.2008.672.675
URL: https://scialert.net/abstract/?doi=pjbs.2008.672.675
INTRODUCTION
Molybdate reduction to molybdenum blue is a phenomenon that is more than one hundred years old. According to Levine (1925), the phenomenon was first reported in E. coli by Capaldi and Proskauer (1896). Since then, other reports on molybdate reduction by other bacterium has been reported in approximately in a ten-year interval (Jan, 1939; Marchal and Girard, 1948; Woolfolk and Whiteley, 1962; Bautista and Alexander, 1972). After a silence of nearly 13 years, microbial molybdate reduction resurfaced again in a report on its reduction by E. coli K12 (Campbell et al., 1985). Sugio et al. (1987, 1988) reported the reduction of molybdate into molybdenum blue by Thiobacillus ferreoxidans and identified the sulfur:ferric ion oxidoreductase (SFORase) as the enzyme responsible for the reduction. Further studies by Yong et al. (1997) however, showed that molybdate reduction in T. ferreoxidans is chemically mediated by ferrous iron added into the media; a known fact of molybdate chemistry (Lee, 1977; Sidgwick, 1984). The efforts of Sugio et al. (1988) were followed by Ghani et al. (1993), who reported that another heterotrophic bacterium, Enterobacter cloacae strain 48 (EC 48) was able to reduce molybdate to molybdenum blue. A new mechanism of molybdate reduction in EC 48 was proposed involving phosphomolybdate as an intermediate between molybdate and molybdenum blue (Shukor et al., 2000). It was also shown that unlike Thiobacillus ferreoxidans where the reduction of molybdate to molybdenum blue is probably due to ferrous iron, in EC 48 it is predominantly enzymatic (Shukor et al., 2002) using the modified method of Munch and Ottow (1983). Using phosphomolybdate as a substrate, the Mo-reducing enzyme was partially purified and characterized (Shukor et al., 2003).
Previously, we have devised a standard curve to quantify molybdenum blue from EC 48 using 12 MP reduced by ascorbate to molybdenum blue (Shukor et al., 2000). This method could be a testing tool to simulate the effect of various interfering substances (inhibitors or activators) on the production of molybdenum blue from E. cloacae strain 48 and for other biological reduction of molybdate to molybdenum blue. This would prevent misleading analysis of the effect of activators and inhibitors to the activity of the molybdenum-reducing microbes in the future.
MATERIALS AND METHODS
Enterobacter cloacae Strain 48 was originally isolated from Chengkau, Malaysia (Ghani et al., 1993) and was grown on agar plate and in low phosphate (2.9 mM phosphate) media (pH 7.0) containing glucose (1%), (NH4)2SO4 (0.3%), MgSO4.7H2O (0.05%), NaCl (0.5%), yeast extract (0.0.5%), Na2MoO4.2H2O (0.242%) and Na2HPO4 (0.05%). All chemicals are of analytical grade. Molybdenum blue is produced in this media but not at high phosphate media (100 mM phosphate). The only difference between the high and low phosphate media is the phosphate concentration.
Preparation of ascorbate-reduced molybdenum blue: Briefly, 12-phosphorous molybdate or phosphomolybdate (BDH) was prepared in distilled water as a 5 mM stock solution and the pH adjusted to pH 5.0 with HCl. Ascorbic acid was prepared as a 25% solution in distilled water and kept at 4°C for a maximum of one week. One hundred microlitres from the 12 MP stock solutions was added to 100 µL ascorbic acid. Suitable volumes of chemicals (phosphate, mercury and arsenate) from stock solutions were added and the final volume adjusted to one milliliter with distilled water. After 12 h of incubation, the absorbance was read at the wavelength of 865 nm.
Preparation of cellular- reduced molybdenum blue: EC 48 culture grown in 250 mL high phosphate media (100 mM phosphate) was used in the inhibitor studies for the cellular reduction of phosphomolybdate since it contained high molybdenum-reducing activity although the media did not turn blue. Cells were harvested after 24 h through centrifugation at 10, 000 g. The pellet was washed with distilled water and resuspended in 100 mL low phosphate media minus yeast extract and 10 mL aliquots was added into sterile bijou bottles. Incubation period was for 24 h. Phosphate (Na2HPO4.2H2O) was added into the bottles from a 1 M stock solution (pH 7.0) at a final concentration from 10 to 200 mM. Heavy metals such as mercury (HgCl2.2H2O, JT Baker, Phillipsburg, USA) and arsenate (AsHNa2O4.7H2O, Fluka, USA) were added into reaction vessels in the same manner as phosphate above. After incubation period has elapsed, the wavelength at OD 865 nm was read. Concentration of inhibitor causing 50% reduction in activity (IC50) was calculated using a one-phase exponential decay model for non-linear regression analysis performed using GraphPad Prism version 4.00 for Windows, GraphPad Software, San Diego California USA, http://www.graphpad.com. The experiments above were carried out in triplicate.
RESULTS AND DISCUSSION
When phosphate was added into EC 48 media, molybdenum blue production from EC 48 decreased significantly. A similar profile was obtained with the ascorbate reduced 12 MP (Fig. 1). The inhibitory effect on the molybdenum reduction in EC 48 is probably due to either the 12 MP complex not forming since molybdate is
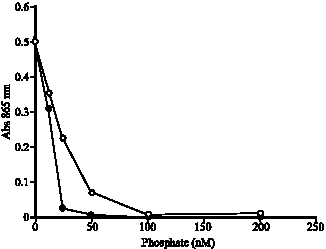 | |
| Fig. 1: | Profile of ascorbate (chemical) reduction of 12-MP (•) and EC 48 (o) reduction of molybdate in the presence of phosphate. The error bars represent mean±standard error of the mean for three replicates |
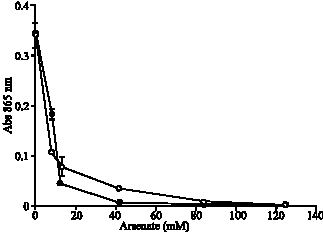 | |
| Fig. 2: | Profile of (chemical) reduction of 12 MP (•) and EC 48 (o) of molybdate in the presence of arsenate. The error bars represent mean±standard error of the mean for three replicates |
not converted to the polyions form at neutral pH (pH 7) or (and) through the disruption of the phosphomolybdate complex at high concentration of phosphate ions. Both conditions could occur simultaneously as observed by Glenn and Crane (1956) who reported that molybdenum blue is unstable in neutral pH and high concentration of phosphate (100 mM). Campbell et al. (1985) also reported that cellular reduction of phosphomolybdate by E. coli K-12 requires an acidic pH (pH 5-6) and high concentration of phosphate exceeding 50 mM is inhibitory to molybdate reduction.
Figure 2 shows that arsenate appears to inhibit Mo-reducing enzyme activity. Arsenate is an inhibitor to oxidative and substrate level phosphorylation
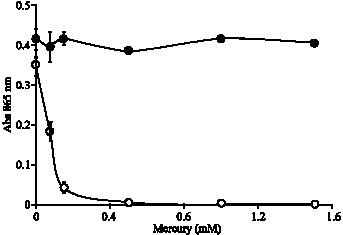 | |
| Fig. 3: | Profile of (chemical) reduction of 12 MP (•) and EC 48 (o) reduction of molybdate in the presence of mercury. The error bars represent mean±standard error of the mean for three replicates |
(Dawson et al., 1969) and thus it is tempting to conclude that arsenate is a true cellular Mo-reducing enzyme inhibitor with a calculated IC50 value of 4.67 mM (R2 = 0.99). However, arsenate also inhibits chemical molybdate reduction by ascorbate with a calculated IC50 value of 6.27 mM (R2 = 0.96). Arsenate is similar to phosphate in terms of forming oxyanions and it was reported that arsenate can replace phosphate ions in the phosphomolybdate complex and would interfere with the phosphate determination method by negatively affecting the Mo-blue synthesis (Clesceri et al., 1989). Thus it can be suggested that the mechanism of inhibition of molybdate reduction by arsenate is at the level of the phosphomolybdate complex by interrupting phosphomolybdate formation or destabilising the complex, both causing a reduction of phosphomolybdate species available for reduction by bacteria as well as chemical reducing agents. Thus care must be exercised in interpreting inhibition results concerning Mo-reducing enzyme.
Mercury significantly reduces bacterial Mo-reducing capability (50%) at the concentration of 0.1 mM (Fig. 3). The calculated IC50 value for mercury is 0.065 mM (R2 = 0.98). Mercury does not affect the ascorbic acid reduction of phosphomolybdate into Mo-blue as arsenate and phosphate did when it was subjected to the experiment of the effect of interfering agents as secondary reactions. This suggests that the inhibition seen with mercury is on the Mo-reducing enzyme. Thus, amongst all of inhibitors tested, only mercury gives a true enzyme inhibition results. The mechanism of mercury inhibition on Mo-reducing enzyme is probably via its binding to the sulfhydryl active sites, inactivating the enzymes (Dawson et al., 1969).
REFERENCES
- Bautista, E.M. and M. Alexander, 1972. Reduction of inorganic compounds by soil microorganisms. Soil Sci. Soc. Am. Proc., 36: 918-920.
CrossRef - Campbell, M.A., A. del Campillo-Campbell and D.B. Villaret, 1985. Molybdate reduction by Escherichia coli K-12 and its chl mutants. Proc. Nat. Acad. Sci. USA., 82: 227-231.
Direct Link - Ghani, B., M. Takai, N.Z. Hisham, N. Kishimoto, M.I.A. Ismail, T. Tano and T. Sugio, 1993. Isolation and characterization of a Mo6+-reducing bacterium. Applied Environ. Microbiol., 59: 1176-1180.
Direct Link - Glenn, J.L. and F.L. Crane, 1956. Studies on metalloflavoproteins: V. The action of silicomolybdate in the reduction of cytochrome c by aldehyde oxidase. Biochim. Biophys. Acta, 22: 111-115.
CrossRefDirect Link - Levine, V.E., 1925. The reducing properties of microorganisms with. special reference to selenium compounds. J. Bacteriol., 10: 217-263.
Direct Link - Munch, J.C. and J.C.G. Ottow, 1983. Reductive transformation mechanism of ferric oxides in hydromorphic soils. Ecol. Bull., 35: 383-394.
Direct Link - Sugio, T., W. Mizunashi, K. Inagaki and T. Tano, 1987. Purification and some properties of sulfur: Ferric-ion oxidoreductase from Thiobacillus ferreoxidans. J. Bacteriol., 169: 4916-4922.
Direct Link - Sugio, T., Y. Tsujita, T. Katagiri, K. Inagaki and T. Tano, 1988. Reduction of Mo6+ with elemental sulfur by Thiobacillus ferreoxidans. J. Bacteriol., 170: 5956-5959.
Direct Link - Woolfolk, C.A. and H.R. Whiteley, 1962. Reduction of inorganic compounds with molecular hydrogen by Micrococcus lactilyticus. I. Stoichiometry with compounds of arsenic, selenium, tellurium, transition and other elements. J. Bacteriol., 84: 647-658.
Direct Link