
Abbasi Fatemeh
Department of Biology, University of Alzahra, Tehran, Iran
Ghafori Sanaz
Department of Marine Biology, Islamic Azad University, Tehran, Iran
Jamili Shahla
Department of Marine Biology, Islamic Azad University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 4 | Page No.: 623-627







ABSTRACT
Stizostedion lucioperca aquaculture/stoking, remains a restrained industry due to several factors such as the paucity of freshwater resources and studies on the physiological responses of this species under environmental changes. The fish were subjected to handling stress by holding them out of the water in a hand-held dip net for 30 sec and netting the fish from the rearing tanks and transferring them to a small confinement tank. Sufficient aeration was supplied to the confinement tank to revert additional stress from oxygen depletion. Then measured changes in plasma cortisol levels and the growth ability (body composition) in Stizostedion lucioperca subjected to handling stress. Blood samples were collected from the fish after exposure to the handling stress. Crude protein (Nx6.25) was determined according to the Kjeldahl method, moisture content was determined by oven drying at 105 ± 2 °C to constant weight and ash by heating in a muffle furnace at 550 °C to constant weight. Total lipids were extracted according to the Bligh and Dyer method. The results indicated that, handling stress significantly increased the plasma levels of cortisol 59.04 ng mL-1 versus 40.83 ng mL-1 in control group. Also the decrease of the level of protein and lipid concentrations show a significant difference between treatment and control (p<0.05). As protein and lipid decreased, moisture increased from 78.19% in control to 80.40% in treatment groups. According to the results, there was no significant change in ash content in control and treatment groups which was about 9%. In other words, it could be emphasized that nutrition-related behavior of Stizostedion lucioperca resulting from the activation of the hypothalamic/inter-renal axis in response to stress despite of different reactions bear resemblance to that of other fishes. Present data indicate that cortisol appears to be adequate to assess stress in Stizostedion lucioperca.
PDF Abstract XML References Citation
How to cite this article
Abbasi Fatemeh, Ghafori Sanaz and Jamili Shahla, 2008. Plasma Cortisol Changes and Body Composition in
Stizostedion lucioperca Exposed to Handling Stress. Pakistan Journal of Biological Sciences, 11: 623-627.
DOI: 10.3923/pjbs.2008.623.627
URL: https://scialert.net/abstract/?doi=pjbs.2008.623.627
DOI: 10.3923/pjbs.2008.623.627
URL: https://scialert.net/abstract/?doi=pjbs.2008.623.627
INTRODUCTION
tress in teleosts is characterized by the immediate release of catecholamines and cortisol; both hormones are concerned with energy reallocation from anabolic activities such as growth toward activities to restore homeostasis (Wendelaar Bonga, 1997). Fish are sensitive to acute and chronic environmental changes and show a stress response (Wedemeyer et al., 1990; Barton and Iwama, 1991; Iwama et al., 1995; Wendelaar Bonga, 1997; Barton et al., 1997; Barton, 2000). In aquaculture, fish are frequently exposed to stressful situations such as handling and confinement (Van der Salm et al., 2006). Acute stresses from different origins are reported to change the hematological parameters in different species (Demers and Bayne, 1997; Jeney et al., 1997; Barcellos et al., 1999; Kubokawa et al., 1999; Lohner et al., 2001; Biswas et al., 2004; Geslin and Auperin, 2004; Wang et al., 2004). Exposure of fish to stressors can elicit physiological changes at multiple levels of animal organization, these alterations are collectively known as the stress response. The initial response to stress represents the perception of an altered state and is characterized by a neuroendocrine response, which includes the release of stress hormones, such as cortisol (Hosoya et al., 2007). Cortisol is the major corticosteroid produced during stress-induced activation of the Hypothalamic-Pituitary-Interrenal (HPI) axis and is considered a principal component of the primary stress response (Donaldson, 1981) cortisol is a major stress-related hormone and its plasma level increases in response to stress (Billard and Gillet, 1981; Pickering et al., 1982; Sumpter and Donaldson, 1986).
The purpose of this study was to investigate the effects of an acute handling and confinement stress on circulating levels of cortisol in Stizostedion lucioperca. In addition, determine protein, lipid, moisture and ash concentration and physiological measurements were made to verify and expand upon.
MATERIALS AND METHODS
Fish: There were two experimental groups: control and stressed fishes (two replicates in each). Twenty eight specimens of Stizostedion lucioperca were obtained from the volga river and transported to one of the culture ponds of the Research Institute of Aquaculture of the Shahid Rajaii, Rasht City, Iran, were used for the stress experiment. Their body weights (means±SEM) were 750±47 g, respectively.
Experiment 1: Acute handling and confinement stress: After one week of acclimation, a group of fourteen was subjected to acute stress according to Biswas et al. (2004). After 48 h, Approximately 1 mL of blood was collected from the caudal vein using a heparinized syringe equipped with a 25 G needle and maintained on ice until centrifugation. Anaesthesia, measurement and blood withdrawal took less than 3 min for five fish. Plasma was separated by 3000 rpm centrifugation at 4°C for 15 min and stored at -80°C for subsequent analysis of different parameters.
Radioimmunoassays: Concentrations of cortisol in the plasma samples were determined by radioimmunoassays as described by Takahashi et al. (1985) and v for cortisol. All samples were measured in duplicate determinations.
Proximate analysis of muscle tissue: Crude protein (Nx6.25) was determined according to the Kjeldahl method (AOAC, 1996), moisture content was determined by oven drying at 105±2°C to constant weight and ash by heating in a muffle furnace at 550°C to constant weight. Total lipids were extracted according to the Bligh and Dyer method (1959), evaporated, weighted and then redissolved in chloroform methanol 9:1, v/v; finally they were stored at a temperature of 0°C.
Statistical analyses: The data were subjected to one -way ANOVA to test the effects of feeding frequency and dietary moisture content. Where significant p<0.05 differences were found in the one-way ANOVA test.
RESULTS AND DISCUSSION
Plasma cortisol levels in fish exposed to acute handling and confinement stress rapidly to maximum value and circulating levels of cortisol were significantly (p<0.05) different between treatments at the time of control (from 40.82 ng mL-1 in control group to 59.04 ng mL-1 in treatment group) (Fig. 1).
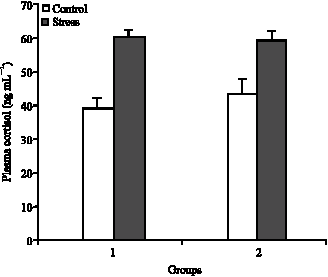 | |
| Fig. 1: | Changes in mean plasma cortisol levels with the standard error (a vertical line on each column) in Stizostedion lucioperca exposed to the acute stress during and after 48 h **: Significantly different at p<0.05 |
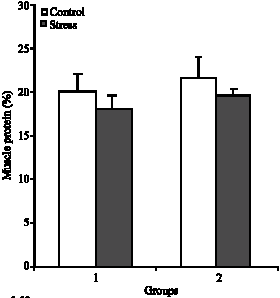
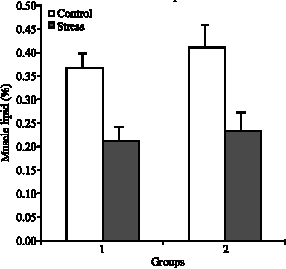
Protein and lipid concentrations show significantly decrease between treatment and control (p<0.05). Protein from 20.82% in control to 18.74% in treatment and lipid from 39% in control to 22% in treatment decreased. Moisture from 78.19% in control to 80.40% in treatment increased (Fig. 2).
Characteristic cortisol elevations of fishes in response to acute stressors tend to range within about 30 and 300 ng mL-1 (Wedemeyer et al., 1990; Barton and Iwama, 1991) but there are notable exceptions. Barton et al. (1997) and Barton (2000) observed that peak levels in cortisol following an acute handling stressor were low in scaphirhynchid sturgeons (Scaphirhynchus sp.) and paddlefish (Polyodon spathula). Their results suggest a trend toward lower stress responses in those chondrosteans compared with teleosts.
Response differences to stressors are clearly evident among closely related fish species and such differences appear to be consistent. Barton (2000) showed that brown trout (Salmo trutta) exhibited greater cortisol increases after brief handling and short-term confinement, respectively, than did rainbow trout (Oncorhynchus mykiss). This difference was also consistent with glucose responses between these two species. Similarly, Barton (2000) found that lake trout (Salvelinus namaycush) were more sensitive to a transport stressor than brook trout (Salvelinus fontinalis), a closely related char species.
The acute handling and confinement stress used in these experiments evoked a clear and similar stress response in Stizostedion lucioperca, confirming and
 | |
 | |
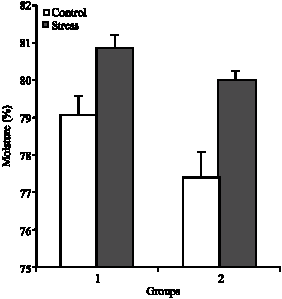 | |
| Fig. 2: | Changes in mean SE protein, lipid and moisture concentration with the standard error (a vertical line on each column) in Stizostedion lucioperca exposed to the acute stress during and after 48 h **: Significantly different at p<0.05 |
expanding upon the results of Biron and Benfey (1994). This includes rapid elevation of plasma cortisol levels, indicative of activation of the HPI axis and increased plasma glucose levels, due to stress-induced mobilization of energy reserves.
The present results in the stress experiment clearly show that Stizostedion lucioperca responded to the acute stress caused by capture, confinement by increasing plasma cortisol levels. Similar results have been reported already in many teleost species including diploid rainbow trout (Dick and Dixon, 1985; Angelidis et al., 1987; Pickering et al., 1987), brown trout (Salmo trutta) (Pickering et al., 1982, 1987), Atlantic salmon (S. salar) (Pickering et al., 1987) and salmon (Fagerlund, 1967; Billard and Gillet, 1981; Pickering et al., 1982; Pickering and Pottinger, 1987; Carragher et al., 1989; Carragher and Sumpter, 1990; Foo and Lam, 1993; Chopin et al., 1996; Jardine et al., 1996; Waring et al., 1996; Clearwater and Pankhurst, 1997).
Present results show that Stizostedion lucioperca responded to acute stress by increasing cortisol. As for the mechanism that determines the maximal level of cortisol in plasma, we suspect the presence of a negative feedback mechanism in the hypothalamus-pituitary-interrenal axes. Suppression of ACTH secretion by elevated cortisol level might determine the maximal level of cortisol as suggested by Pickering and Pottinger (1987).
The cortisol in plasma in Stizostedion lucioperca captured in the lake were measured to confirm whether the cultured Stizostedion lucioperca used in the stress experiment are physiologically equivalent to wild individuals. The values of the physiological parameters observed at the beginning of the experiment in cultured fish were comparable to the values observed in wild individuals captured. Accordingly, the initial condition of the cultured fish can be regarded as equivalent to the wild fish after ovulation but before spawning under the present experimental conditions. This allows us to further generalize the results of the stress experiment.
In the present study, transporting the fish, keeping the fish in the pond and handling the fish for bleeding in all likelihood caused stress and affected plasma hormone and body composition.
tudies of protein concentration of control group in autumn and winter show that protein concentration is increased (from 20.82% in control to 18.74% in treatment) which is due to growth of physiology and tissue substitution against to moisture. Therefore structural role and shaping body tissue by protein is important. In this species, increase in cortisol level causes increased in lipid metabolism, lipid decomposed and therefore a release of FFA. Increasing oxidation of lipid acids lead to intensification of metabolite. Sadovy (1993), also has studied about body composition in different phase of reproductive and found that spawning decrease tissue reserve such as lipid. Steven et al. (2000) have studied body composition biochemistry in the season changes as a stress stimulus in fish. In this research we found that lipid in control group 39% is decreased to 22% in treatment group. That is statistically significant. These results consist of data and the difference is due to cortisol role as a lipitic factor in the treatment. As protein and lipid decreased, moisture from 78.19% in control to 80.40% in treatment increased. Therefore protein and lipid tissue substitution with moisture tissue increase. Moisture group in autumn and winter shows that, in winter is decreased in control. According to the results, there is no significant change in ash (control and treatment) it is about 9%.
However primary and secondary physiological response to season changing stress in Stizostedion lucioperca is completely normal, but this species is as an active and hunter species tertiary response (low active changing behavior).
In conclusion, present results have shown that Stizostedion lucioperca exhibit the typical physiological responses to acute stress induced by handling and brief confinement and acute stress increased the plasma levels of cortisol in this species. Similarly, the acute stress induced decrease in total protein level parallels the findings in other species.
REFERENCES
- Angelidis, P., F. Baudin-Laurencin and P. Youinou, 1987. Stress in rainbow trout, Salmo gairdneri: Effects upon phagocyte chemiluminescence, circulating leucocytes and susceptibility to Aeromonas salmonicida. J. Fish Biol., 31: 113-122.
CrossRef - Barcellos, L.J.G., S. Nicolaiewsky, S.M.G. de Souza and F. Lulhier, 1999. Plasmatic levels of cortisol in the response to acute stress in Nile tilapia, Oreochromis niloticus (L.), previously exposed to chronic stress. Aquacult. Res., 30: 437-444.
CrossRefDirect Link - Barton, B.A. and G.K. Iwama, 1991. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish Dis., 1: 3-26.
CrossRefDirect Link - Barton, B.A., 2000. Salmonid fishes differ in their cortisol and glucose responses to handling and transport stress. North Am. J. Aquacult., 62: 12-18.
Direct Link - Biswas, A.K., M. Maita, G. Yoshizaki and T. Takeuchi, 2004. Physiological responses in Nile tilapia exposed to different photoperiod regimes. J. Fish Biol., 65: 811-821.
Direct Link - Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Demers, N.E. and C.J. Bayne, 1997. The immediate effects of stress on hormones and plasma lysozyme in rainbow trout. Dev. Comp. Immunol., 21: 363-373.
CrossRefPubMedDirect Link - Dick, P.T. and D.G. Dixon, 1985. Changes in circulating blood cell levels of rainbow trout, Salmo gairdneri Richardson, following acute and chronic exposure to copper. J. Fish Biol., 26: 475-481.
CrossRefDirect Link - Geslin, M. and B. Auperin, 2004. Relationship between changes in mRNAs of the genes encoding steroidogenic acute regulatory protein and P450 cholesterol side chain cleavage in head kidney and plasma levels of cortisol in response to different kinds of acutestress in the rainbow trout (Oncorhynchus mykiss). Gen. Comp. Endocrinol., 135: 70-80.
Direct Link - Hosoya, S., S.C. Johnson, G.K. Iwama, A.K. Gamperl and L.O.B. Afonso, 2007. Changes in free and total plasma cortisol levels in juvenile haddock (Melanogrammus aeglefinus) exposed to long-term handling stress. Comp. Biochem. Physiol. Part A: Mol. Integrative Physiol., 146: 78-86.
Direct Link - Iwama, G.K., J.D. Morgan and B.A. Barton, 1995. Simple field methods for monitoring stress and general condition of fish. Aquacult. Res., 26: 273-282.
Direct Link - Jeney, G., M. Galeotti, D. Volpatti, Z. Jeney and D.P. Anderson, 1997. Prevention of stress in rainbow trout (Oncorhynchus mykiss) fed diets containing different doses of glucan. Aquaculture, 154: 1-15.
CrossRefDirect Link - Kubukawa, K., T. Watanabe, M. Yoshizaki and M. Iwata, 1999. Effects of acute stress on plasma cortisol, sex steroid hormone and glucose leveles in male and female sockeye salmon during the breeding season. Aquaculture, 172: 335-349.
Direct Link - Lohner, T.W., R.J. Reash, V.E. Willet and J. Fletcher, 2001. Assessment of tolerant sunfish populations (Lepomis sp.) inhabiting selenium-laden coal ash effluents: 3. serum chemistry and fish health indicators. Ecotoxicol. Environ. Safety, 50: 225-232.
CrossRef - Pickering, A.D., T.G. Pottinger and P. Christie, 1982. Recovery of the brown trout, Salmo trutta, from acute handling stress: A time course study. J. Fish. Biol., 20: 229-244.
CrossRef - Sadovy, Y., 1993. Size, composition and spatial structure of the annual spawning aggregation of the red hind, Epinephelus guttatus. Pisces: Serranidae, 2: 399-406.
Direct Link - Craig, S.R., D.S. MacKenzie, G. Jones and D.M. Gatlin, 2000. Seasonal changes in the reproductive condition and body composition of free-ranging red drum, Sciaenops ocellatus. Aquaculture, 190: 89-102.
CrossRef - Van der Salm, A.L., M. Pavlidis, G. Flik and S.E. Wendelaar Bonga, 2006. The acute stress response of red porgy, Pagrus pagrus, kept on a red or white background. Gen. Comp. Endocrinol., 145: 247-253.
CrossRef - Wang, C., V.W. King and L.C. Woods III, 2004. Physiological indicators of divergent stress responsiveness in male striped bass broodstock. Aquaculture, 232: 665-678.
CrossRef - Waring, C.P., R.M. Stagg and M.G. Poxton, 1996. Physiological responses to handling in the turbot. J. Fish Biol., 48: 161-173.
CrossRefDirect Link - Bonga, S.E.W., 1997. The stress response in fish. Physiol. Rev., 77: 591-625.
CrossRefPubMedDirect Link