
R. Sarnthima
Protein and Enzyme Technology Research Unit, Department of Chemistry, Faculty of Science, Mahasarakham University, Maha Sarakham 44150, Thailand
S. Khammuang
Protein and Enzyme Technology Research Unit, Department of Chemistry, Faculty of Science, Mahasarakham University, Maha Sarakham 44150, Thailand
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 62-67







ABSTRACT
The extracellular enzymes from Pleurotus sajor-caju were studied for lignin degrading enzyme patterns and dye decolourisation potential. Laccases are major ligninolytic enzymes excreted by the fungus. The results from a native-PAGE revealed that there were at least two isoenzymes. The crude enzyme had a pH and a temperature optimum at 6.0 and 40°C, respectively when 2,2'-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) was used as substrate. The pH and thermal stability were at 5.0 and 30°C. The pH optima for decolourisation of Indigo Carmine and Methyl Red were at 5.0 and 6.0, respectively. Indigo Carmine could be decolorized efficiently above 90% within 180 min, whereas Methyl Red could be decolorized only 3.5%. High efficiency decolourisation of Indigo Carmine makes this fungus to be a promise choice in biological treatment of waste water containing Indigo Carmine.
PDF Abstract XML References Citation
How to cite this article
R. Sarnthima and S. Khammuang, 2008. Evaluation of Dyes Decolourisation by the Crude Enzyme from Pleurotus sajor-caju Grown on Sorghum Seed Media. Pakistan Journal of Biological Sciences, 11: 62-67.
DOI: 10.3923/pjbs.2008.62.67
URL: https://scialert.net/abstract/?doi=pjbs.2008.62.67
DOI: 10.3923/pjbs.2008.62.67
URL: https://scialert.net/abstract/?doi=pjbs.2008.62.67
INTRODUCTION
Wood-rotting fungi, characterized by their ability to degrade lignin and cellulose, are predominant microorganisms of wood degradation. Some wood-rotting fungi, especially white-rot fungi, appear to have some potential for bioremediation applications due to their non-specific system which they have developed for depolymerization and mineralization of the complex and recalcitrant polymer of lignin (Hatakka, 1994). As the lignin degradation system of these fungi is not very substrate specific, they are able to transform and sometimes completely mineralize a variety of persistent environmental pollutants (Cameron et al., 2000). Among such compounds there are many synthetic dyes characterized by high stability in light and during washing, which give them recalcitrance to biodegradation (Gramss et al., 1999). Between 10 and 15% of the total dyes consumed in dyeing processes may be found in wastewater. Most of these compounds are highly resistant to microbial attack. Therefore, it is hard to remove them from effluents by means of conventional biological wastewater treatments.
The degradation of dyestuffs by fungi has been carried out with either whole cultures or crude enzyme preparations of extracellular ligninolytic enzymes. Decolourisation of azo, anthraquinonic, heterocyclic, triphenylmethane and polymeric dyes and their partial mineralization by enzymatic and non-enzymatic systems of these fungi have been reported (Ferreira et al., 2000; Swamy and Ramsay, 1999). Several numbers of white-rot fungi have been reported to produce the lignin-degrading enzymes laccase, Lignin Peroxidases (LiP) and Manganese Peroxidases (MnP), or at least one of these enzymes (Eggert et al., 1996).
Laccases (benzenediol: oxygen oxidoreductase, EC 1.10.3.2) have very broad substrate specificity and they catalyze the removal of a hydrogen atom from the hydroxyl group of ortho and para-substituted mono and polyphenolic substrates and from aromatic amines by one-electron abstraction to form free radicals capable of undergoing further reactions such as depolymerization, repolymerization, demethylation, or quinone formation (Thurston, 1994). The rather broad substrate specificity of laccases may be additionally expanded by addition of redox mediators, such as ABTS, 1-hydroxybenzotriazole (HBT) Claus et al., 2002). Laccases from several fungi have been reported that can be used for the treatment of effluents containing synthetic recalcitrant dyes. T. hirsute and a purified laccase were able to degrade triarylmethane, indigoid, azo and anthraquinone dyes used in dyeing textiles (Abadulla et al., 2000) as well as 23 industrial dyes (odriguez et al., 1999).
Pleurotus sajor-caju is one of popular commercial mushrooms grown in several parts of Thailand and other parts worldwide. Four laccase isozyme genes, Psc lac1, 2, 3 and 4 have been cloned from this mushroom (Soden and Dobson, 2001). Apart from laccases, the mushroom excreted ligninolytic enzymes MnP (Murugesan et al., 2006). The Psc lac4 gene from P. sajor-caju has been cloned and expressed in the heterologous host Pichia pastoris, purified and biochemical characterized (Soden et al., 2002). Moreover, an extracellular laccase was isolated and purified from this fungus grown in submerged culture in a bioreactor and used to investigate its ability to decolorize three azo dyes (Murugesan et al., 2006).
In the North Eastern area of Thailand, one of small popular enterprises is silk. Many synthetic dyes have been used in silk dyeing even though natural ones is considerate to alternatively use nowadays; of course some are discarded into the environment. Therefore, in environmental concerns it would be very valuable as if we could develop an easy and uncomplicated way for dye-containing wastewater treatment. Due to availability of this edible mushroom, it is made us interested to evaluate the potential of dye decolourisation by its enzymes. In this present work, we have studied the ligninolytic enzyme pattern, some characteristics of the crude enzyme from P. sajor-caju and its potential to decolorize two structurally different dyes.
MATERIALS AND METHODS
Microorganism: The white-rot fungus P. sajor-caju was obtained from the commercial mushroom farm in Ka La Sin province (Northeast, Thailand). The stock cultures were maintained on potato dextrose agar medium and stored at 4°C with periodic subculture. All experiments were performed at the Protein and Enzyme Technology Research Unit, Faculty of Science, Mahasarakham University during August 2006 and February 2007.
Enzyme extraction, activity assays and characterization: The fungus P. sajor-caju was grown on sorghum seed media without any mineral supplementation for about 2 weeks at room temperature. The crude enzyme was extracted with 50 mM sodium acetate buffer (pH 4.5), filtered through sheet cloth to remove solid stuffs and then subjected to a centrifugation at 3,000 rpm for 15 min to discard the precipitate. The clear supernatant, crude enzyme was concentrated by ammonium sulfate precipitation at 0-80% saturation. After centrifugation at 10,000 rpm, 15 min at 4°C, the precipitate was re-suspended in 50 mM sodium acetate buffer (pH 4.5) prior to enzyme activity assays and protein determination.
Laccase (EC 1.10.3.2) activity was spectrophotometric assayed at 32°C of oxidized ABTS (blue green color cation radical) as previously described (Khammuang and Sarnthima, 2007). Briefly, the assay mixture contained 0.1 mM ABTS and 0.1 M sodium acetate buffer (pH 4.5). Oxidation of ABTS was followed by absorbance increase at 420 nm (ε = 36,000 M-1 cm-1).
Lignin peroxidase (EC 1.11.1.14, LiP) activity was measured according to Collins et al. (1996) using veratryl alcohol as substrate in the presence of H2O2.
Manganese peroxidase (EC 1.11.1.13, MnP) activity was measured the oxidation of MBTH (3-methyl-2-benzothiazolinone hydrazone) and DMAB (3, 3-dimethylaminobenzoic acid) in the presence of H2O2 , a modified method of Ngo and Lenhoff (1980). The results were corrected by activities in the reactions without manganese (Mn-independent peroxidase, MnIP) where manganese sulfate was substituted by ethylenediaminetetraacetate (EDTA) to chelate Mn present in the samples. All assays were performed in duplicate, with an average sample mean deviation of less than 10%. One unit (U) of enzyme activity was defined as the amount of enzyme that catalyzed the formation of 1.0 μmol of product per minute.
Estimation of the laccase optimum pH was measured with ABTS substrate in 50 mM sodium citrate-phosphate buffer (pH 2.0-8.0). The optimum temperature was determined in the range of 30-60°C. The pH stability was tested in the ranges of pH 3.0-8.0 while temperature stability was performed at 30.0-60.0°C based on ABTS assay.
Protein assays: Protein concentration was determined by the Bio-Rad Protein Assay Reagent (Bio-Rad) with bovine serum albumin as a standard.
Decolourisation experiments: Two dyes were tested for decolourisation at a wavelength with maximum absorbance of each dye (610 nm for Indigo Carmine and 524 nm for Methyl Red). Indigo Carmine concentrations were used at 50-500 μM while Methyl Red concentration used from 5,000-50,000 μM. The enzyme activity in the reaction mixture was 1 U per 5.0x10-8 mole of dye. Experiments were monitored immediately using a JENWAY 6400 Spectrophotometer (LABQUIB, England) after enzyme addition and periodically time interval. Dye decolourisation was expressed in terms of percentage calculated according to the following equation.
| Where: | ||
| A0 | = | An absorbance at λmax of each dye immediately measured after enzyme addition. |
At | = | An absorbance at λmax of each dye after each time intervals. |
RESULTS
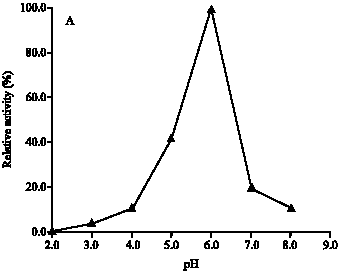
Enzyme extraction and characterization: P. sajor-caju grown on sorghum seed media with no mineral supplementation excreted extracellular ligninolytic enzymes mainly laccase (3.340 U mL-1), some of Mn-independent peroxidase, MnIP (0.217 U mL-1) and very small amount of LiP (0.030 U mL-1). The laccase activity from this fungus showed at least two isoenzymes with difference in electric mobility as a result from a native- PAGE (Fig. 1A). The crude enzyme when using ABTS as substrate, had a sharp pH optimum at 6.0 (Fig. 2A) and the optimal temperature at 40°C (Fig. 2B). At temperature of 60°C, the laccase activity remained approximately 60% compared to those of the optimum temperature.
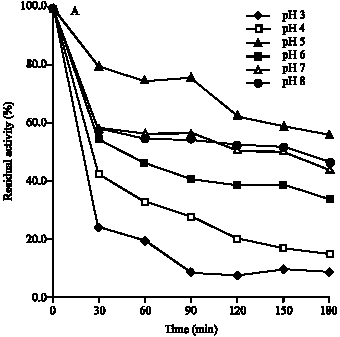
The enzyme stability experiments as shown in Fig. 3A revealed that storage pH had the enormous effect on stability of laccase. It was found that the enzyme kept at 4°C in buffer pH 5.0 retained maximum activity. Within 180 min, the remaining activity was more than 60% from the original. The laccase activity within 180 min was remained less than 10% when kept in the buffer pH 3.0. Moreover, the activity observed less than 20 and 40% when buffer pH 4.0 and 6.0 were used to store, respectively. Neutral (pH 7.0) or basic condition (pH 8.0), the results showed that more than half of activity was lost.
Temperature stability of the crude enzyme was also investigated at the optimum pH condition. Among various temperatures studied, the best condition for enzyme
 |
| Fig. 1: | Native PAGE analysis of the crude enzyme from Pleurotus sajor-caju stained with ABTS substrate (A) and protein bands stained with coomassie brilliant blue R-250 (B) |
stability was at 30°C (Fig. 3B) and at this temperature enzyme activity remained more than 95% after 120 min of incubation. When kept the crude enzyme at 40°C, the laccase activity left above 80%, whereas only about 30% and less than 10% activity remaining when the crude enzyme were kept at 50 and 60°C, respectively.
Effect of pH on dye decolourisation: Each dyes might be susceptible to be efficiently degraded in different pH values, therefore two model dyes with different chemical structures were evaluated for the pH dependent decolourisation by the crude enzyme from P. sajor-caju. The results showed that the optimum pH for Indigo Carmine and Methyl Red decolourisation were shown in Table 1. Indigo Carmine dye could be decolorized at pH 3.0-8.0 with a maximum decolourisation activity at pH 5.0, while Methyl Red was decolorized at pH 4.0-8.0
 |
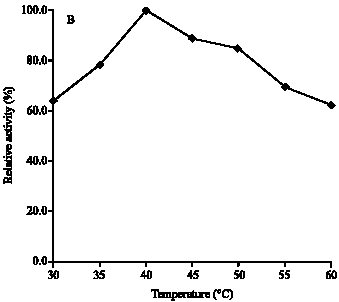 | |
| Fig. 2: | pH optimum condition (A) and temperature optimum (B) of the crude enzyme from Pleurotus sajor-caju according to ABTS assay. Data were averaged from a duplicate experiment |
| Table 1: | Relative decolourisation percentage of each dye at various pH valuesa |
 | |
a: The reactions were performed using 50 mM sodium citrate-phosphate buffer and dye 5.0x10-8 mole/Unit of enzyme at 32°C, 30 min. Data were averaged from a duplicate experiment | |
 |
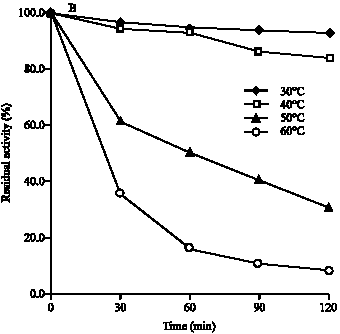 | |
| Fig. 3: | pH stability when kept at 4°C (A) and temperature stability when kept at buffer pH 5.0 (B) of the crude enzyme from Pleurotus sajor-caju. Data were averaged from a duplicate experiment |
with the highest percentage at pH 6.0. Therefore, pH 5.0 and pH 6.0 were chosen for following studies of Indigo Carmine and Methyl Red decolourisations.
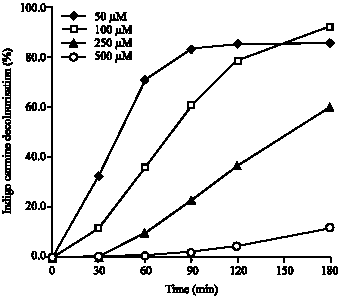 | |
| Fig. 4: | Effect of initial Indigo Carmine concentration on decolourisation ability by the crude enzyme from Pleurotus sajor-caju. The reactions were performed in 50 mM sodium citrate-phosphate buffer, pH 5.0 and 1 Unit of enzyme at 32°C. Data were averaged from a duplicate experiment |
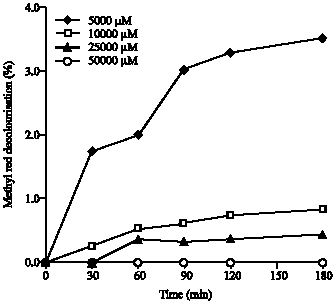 | |
| Fig. 5: | Effect of initial Methyl Red concentration on decolourisation ability by the crude enzyme from Pleurotus sajor-caju. The reactions were performed in 50 mM sodium citrate-phosphate buffer, pH 6.0 and 1 Unit of enzyme at 32°C. Data were averaged from a duplicate experiment |
Effect of initial dye concentration on decolourisation Indigo carmine: The indigoid dye Indigo Carmine decolourisation by crude enzyme from P. sajor-caju at different initial concentrations (50-500 μM) as a function of time (Fig. 4). Within 180 min, at a fixed amount of enzyme it was observed that increasing initial concentration the decolourisation ability was either slower or lower decolourisation levels. At initial dye concentration of 50 μM, more than 80% of dye decolourisation was achieved within 90 min and remained constantly after that. The initial dye concentration that yielded the highest decolourisation percentage was of 100 μM (>90% within 180 min). The initial concentration of dye above this (250 and 500 μM) gave 60 and 10% decolourisation, respectively.
Methyl red: The decolourisation of Methyl Red, an azo dye (Fig. 5) which, unlike Indigo Carmine, decolourisation was very poor at all initial dye concentration tested at optimum pH 6.0. The highest decolourisation percentage, in the range of initial dye concentration tested, only 3.5% was observed within 180 min.
DISCUSSION
White-rot fungus, P. sajor-caju from our experiment grown on sorghum seed media without mineral supplementation showed that there are at least two isoenzymes of laccases. The crude enzyme had optimal activity with ABTS as substrate at 40°C and pH 6.0. These optimal conditions are slightly different from the purified laccase investigated by Murugesan et al. (2006), which reported that the optimal conditions were 40°C and pH 5.0. However, the results from our work exhibited the higher optimum temperature and less acidic pH than those of the purified recombinant Psc lac4 which displayed optimal activity at 35°C and pH 3.5 (Soden et al., 2002). According to these results, we suggest that laccase in our crude enzyme may be the other isoenzymes of P. sajor-caju different from previous reports.
Structurally different dyes were not decolorized to the same extent. In present experiment, the Indigo Carmine, an indigoid dye, was decolorized in a high percentage. At initial dye concentration lower than 100 μM, more than 80% of dye decolourisation was achieved within 90 min. There has been a report that the decolourisation of this dye by laccase from T. villosa was approximately 20% within 1 h (Basto et al., 2007). This was relatively low when compared to the results from present experiments. From the same report, they also observed that the best results (more than 65% of decolourization) were attained when the dye solution was treated with ultrasound and enzyme stabilized with polyvinyl alcohol. It may point out that apart from the unique characteristics of ligninolytic enzymes of different fungal species, laccase alone may or may not be able to completely decolorize the Indigo Carmine solution. This result was in agreement with other reported studies. T. hirsuta immobilized on stainless steel sponge grown in a 1 L bioreactor supplemented with 1 mM copper sulphate, the textile dye Indigo Carmine was almost totally degraded in 3 days (Rodriguez et al., 2004).
The complete decolourisation of Indigo Carmine was only achieved by a laccase from T. hirsuta when more than 30 h of treatment with the enzyme immobilized in alginate beads (Dominguez et al., 2005). Thus in our case, a good Indigo Carmine decolourisation might be involved somehow with other enzymes apart from laccases. Mn-independent peroxidase, another ligninolytic enzyme also found in the crude extract of P. sajor-caju. Even though its activity was approximately 15 times less than laccase activity, it might concert in decolourisation of Indigo Carmine dye somehow. However, the decolourisations of this dye by the purified enzymes are still worth to study in order to more understand its mechanism.
On the other hand, Methyl Red, an azo dye, decolourisation ability by the crude enzyme from P. sajor-caju was very low. A low efficiency in decolourisation of some azo dyes, compared to other dye types, was also reported for Thelephora sp. (Selvam et al., 2003) which showed that a maximum of 19% Orange G was removed by laccase (15 U mL-1) whereas LiP and MnP at the same concentration decolorized 13.5 and 10.8%, respectively. They also reported that a maximum decolourisation of 12.0 and 15.0% for Congo Red and Amido Black 10B, respectively was recorded by the laccase. However, decolourisation by the fungus in the bioreactor provided higher efficiency of dye removal (Selvam et al., 2003; Tavcar et al., 2006). Murugesan et al. (2006) reported that the purified laccase from P. sajor-caju showed high efficiency for decolorized azo dyes such as Acid Red 18 (90%), Acid Black 1 (87%) and Direct Blue 71 (72%) within 24 h incubation with 10-12 U mL-1.
From present results, the crude enzyme from P. sajor-caju showed fairly acidic pH of catalytic activity and fairly high temperature optimum and quite stable at room temperature (30-40°C) as well as its high potential in Indigo Carmine decolourisation, suggest that it could be used for treatment of this type of industrial dye effluents. In addition, other dyes in the same group or other groups of dyes such as anthraquinonic and triphenylmethane dyes should also be further investigated. However, redox mediators might be applied to improve decolourisation of azo dye type. Purification and characterizations of each isoenzyme as well as their dye decolourisation ability are underway in our laboratory.
ACKNOWLEDGMENTS
This research was financially supported by a grant from the Faculty of Science, Mahasarakham University (Fiscal year 2006).
REFERENCES
- Abadulla, E., T. Tzanov, S. Costa, K.H. Robra, P.A. Cavaco and G.M. Guebitz, 2000. Decolourisation and detoxification of textile dyes with a laccase from Trametes hirsute. Applied Environ. Microbiol., 66: 3357-3362.
Direct Link - Basto, C., C.J. Silva, G. Gubitz and A. Cavaco-Paulo, 2007. Stability and decolourisation ability of Trametes villosa laccase in liquid ultrasonic fields. Ultrason. Sonochem., 14: 355-362.
CrossRefDirect Link - Cameron, M.D., S. Timofeevski and S.D. Aust, 2000. Enzymology of Phanerochaete chrysosporium with the respect to the degradation of recalcitrant compounds and xenobiotics. Applied Microbiol. Biotechnol., 54: 751-758.
CrossRefPubMedDirect Link - Claus, H., G. Faber and H. Konig, 2002. Redox-mediated decolorization of synthetic dyes by fungal laccases. Appl. Microbiol. Biotechnol., 59: 672-678.
CrossRefDirect Link - Collins, P.J.J., M.J.J. Kotterman, J.A. Field and D.W. Dobson, 1996. Oxidation of antracene and benzo(a)pyrene by laccase from Trametes versicolor. Applied Environ. Microbiol., 62: 4563-4567.
Direct Link - Dominguez, A., S.R. Couto and M.A. Sanroman, 2005. Dye decolorization by Trametes hirsuta immobilized into alginate beads. World J. Microbiol. Biotechnol., 21: 405-409.
CrossRefDirect Link - Eggert, C., U. Temp and K.E. Eriksson, 1996. Laccase-producing white-rot fungus lacking lignin peroxidase and manganese peroxidase. ACS Symp. Ser., 655: 130-150.
Direct Link - Ferreira, V.S., D.B. Magalhaes, S.H. Kling, J.G. Da Silva and E.P.S. Bon, 2000. N-demethylation of methylene blue by lignin peroxidase from Phanerochaete chrysosporium. Applied Biochem. Biotechnol., 84: 255-265.
Direct Link - Gramss, G., B. Kirsche, K.D. Voigt, T. Gunther and W. Fritsche, 1999. Conversion rates of five polycyclic aromatic hydrocarbons in liquid cultures of fifty-eight fungi and the concomitant production of oxidative enzymes. Mycol. Res., 103: 1009-1018.
CrossRef - Hatakka, A., 1994. Lignin-modifying enzymes from selected white-rot fungi: Production and role from in lignin degradation. FEMS Microbiol. Rev., 13: 125-135.
CrossRefDirect Link - Khammuang, S. and R. Sarnthima, 2007. Laccase from spent mushroom compost of Lentinus polychrous Lev. and its potential for remazol brilliant blue R decolourisation. Biotechnology, 6: 408-413.
CrossRefDirect Link - Murugesan, K., M. Arulmani, I.H. Nam, Y.M. Kim, Y.S. Chang and P.T. Kalaichelvan, 2006. Purification and characterization of laccase produced by a white rot fungus Pleurotus sajor-caju under submerged culture condition and its potential in decolorization of azo dyes. Applied Microbial. Biotechnol., 72: 939-946.
Direct Link - Ngo, T.T. and H.M. Lenhoff, 1980. A sensitive and versatile chromogenic assay for peroxidase and peroxidase-coupled reactions. Anal. Biochem., 105: 389-397.
Direct Link - Rodriguez, E., M.A. Pickard and R. Vazquez-Duhalt, 1999. Industrial dye decolorization by laccases from ligninolytic fungi. Curr. Microbiol., 38: 27-32.
Direct Link - Couto, S.R., M.A. Sanroman, D. Hofer and G.M. Gubitz, 2004. Stainless steel sponge: A novel carrier for the immobilisation of the white-rot fungus Trametes hirsuta for decolourization of textile dyes. Bioresour. Technol., 95: 67-72.
CrossRefDirect Link - Selvam, K., K. Swaminathan and C. Keo-Sang, 2003. Decolourisation of azo dyes and a dye industry effluent by a white rot fungus Thelephora sp. Biores. Technol., 88: 115-119.
CrossRef - Soden, D.M. and A.D. Dobson, 2001. Differential regulation of laccase gene expression in Pleurotus sajor-caju. Microbiology, 147: 1755-1763.
Direct Link - Soden, D.M., J. O'Callaghan and A.D.W. Dobson, 2002. Molecular cloning of a laccase isozyme gene from Pleurotus sajor-caju and expression in the heterologous Pichia pastoris host. Microbiology, 148: 4003-4014.
CrossRefDirect Link - Swamy, J. and J.A. Ramsay, 1999. The evaluation of white rot fungi in the decoloration of textile dyes. Enzyme Microb. Technol., 24: 130-137.
CrossRefDirect Link - Tavcar, M., K. Svobodova, J. Kuplenk, C. Novotny and A. Pavko, 2006. Biodegradation of azo dye RO16 in different reactors by immobilized Irpex lacteus. Acta. Chim. Slov., 53: 338-343.
Direct Link - Thurston, C.F., 1994. The structure and function of fungal laccases. Microbiology, 140: 19-26.
Direct Link