
M. Sedghi
Faculty of Agronomy and Plant Breeding, College of Agriculture, University of Mohaghegh Ardebili, Ardebil, Iran
R. Seyed Sharifi
Faculty of Agronomy and Plant Breeding, College of Agriculture, University of Mohaghegh Ardebili, Ardebil, Iran
A. Gholipouri
Faculty of Agronomy and Plant Breeding, College of Agriculture, University of Mohaghegh Ardebili, Ardebil, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 4 | Page No.: 595-600







ABSTRACT
Determining the effect of different sources of nitrogen and weeds on two varieties of soybean, an experiment was conducted out at research station of Tabriz University, Iran. Treatments were two varieties (Williams and Harcor), four nitrogen levels (two levels of symbiotic bacteria and two levels of urea) and weeds (with and without control). Results showed that three various factors can significantly change the Photosynthetically Active Radiation (PAR) amount at different layers of canopy. Solar radiation decreased in a vertical profile of canopy because of increasing Leaf Area Index (LAI). At the bottom level of canopy only 6% of total arrived PAR was recorded. Increasing LAI caused a sharp decrease in Light Interception Efficiency (LIE) in the canopy. Weeds significantly affected root nodules in dry and fresh weights. So, in weedy condition of soybean canopy, accumulation of dry matter decreases due to decline in LIE. On the other hand, weeds can strongly affect on nodules activity and decrease potential of nitrogen fixation by symbiosis bacteria.
PDF Abstract XML References Citation
How to cite this article
M. Sedghi, R. Seyed Sharifi and A. Gholipouri, 2008. Practical Methods for Increasing Light Interception Efficiency and Root Growth in Soybean. Pakistan Journal of Biological Sciences, 11: 595-600.
DOI: 10.3923/pjbs.2008.595.600
URL: https://scialert.net/abstract/?doi=pjbs.2008.595.600
DOI: 10.3923/pjbs.2008.595.600
URL: https://scialert.net/abstract/?doi=pjbs.2008.595.600
INTRODUCTION
Leaf Area Index (LAI) and architecture of leaves are two main characteristics that define light interception in the canopy (Rao et al., 2002). There is a linear relation between the cumulative dry matter production and cumulative light interception that slop of this line defines Radiation Use Efficiency (RUE) (Purcell et al., 2002). The cumulative amount of light intercepted by a crop depends upon the number of days of light accumulation, the quantity of daily radiation and fraction of light intercepted by the crop on a daily basis. So, plants with long growth season and smooth leaves have greater efficiency in light interception. In addition, planting date and density of plants can alter the intercepted light (Purcell et al., 2002). Soybean is a self-destructive plant because of its grain high requirements to nitrogen. Redistribution of nitrogen from vegetative parts specially leaves to grains causes leaf senescence that can affect light interception by decreasing LAI (Sinclair and de Witt, 1976; Sinclair et al., 2007; Ghosh et al., 2006).
In a weed-crop mixture canopy, competing for light causes yield decrease in crops (Holt, 1995). A crop canopy drastically alters both light intensity and quality. Although canopies provide effective weed suppression, some weeds are adapted to survive in poor light environments. Selection of competitive cultivars and appropriate management techniques will improve weed control through competition for light (Bussan et al., 1997). Weed interference with soybean can significantly decrease leaf area (Van-Acker et al., 1993) and leaf area duration (Graham et al., 1988) that indirectly changes solar intercepted radiation. Soybean is a Nitrogen Fixing Species (NFS) and its fixing efficiency depends upon the bacterial strain, nitrogen availability and soil physiochemical properties (Ofosu Budu et al., 1993; Song et al., 1993; Singleton and Van Kessel, 1987). Root growth and nodulation of roots are related together. N2 fixation creates a strong localized sink for photosynthate that affects both root and nodule development. The control of carbon flow to roots and nodules is affected by the output of N2 fixation products from nodules. The effects of nitrogen assimilation on carbon partitioning to roots are similar whether the source of N is inorganic or from N2 fixation (Singleton and Van Kessel, 1987; Corre-Hellou et al., 2007; Greci et al., 2007).
The objectives of this study were to determine the effects of different sources of nitrogen (symbiotic fixation and urea) and weeds on soybean leaf area index, solar radiation interception in different strata of canopy, root-shoot growth and nodules formation.
MATERIALS AND METHODS
Field experiments were carried out at research station of Tabriz University (46° 17´ E, 38° 5´ N) with mean annual
| Table 1: | Manner of fertilizer (kg ha-1) application in plots |
 | |
| V and R are soybean development stages according to Fehr and Caviness (1977), N is different nitrogen levels | |
rainfall and mean annual air temperature, respectively 271.3 mm and 10°C. The experimental plots were laid out in a factorial experiment with three replications that arranged in a randomized complete block design. Plots had 5 rows with 60 cm row spacing. Treatments were varieties (Williams, maturity group three and Harcor, maturity group two), four N levels (Table 1) and weeds (without control and hand-weeding). Planting dates were 19 May 2004 and 12 May 2005.
To evaluation of dry and fresh weights of nodules, three pots (60x20 cm) were placed in plots with three plants in them. At R2 stage of soybean growth (Fehr and Caviness, 1977), 0.5 m of one row of each plot was harvested to determining leaf area. In this stage, solar radiation was measured with Sunscan using a 1 m line quantum sensor in three different vertical strata: top, middle and bottom layers of canopy. These measurements were done in four sequential days in similar hour of day for each block. Excavation method (Boehm, 1979) with 1 m soil depth was used for main root length measurement.
Data were first tested for normality and with in groups homogenesity (Mstat-C software). Then analysis of variance was done by SAS and combined analysis with Mstat-C. Means were compared with Duncan test at 1% probability level. Regression analysis was conducted out with SPSS13 and graphs were drawn with Excel2003.
RESULTS AND DISCUSSION
Solar radiation changes: Top surface of soybean canopy was the reference for light measurement. Separate and combined analyses of variance for top level of canopy radiation showed significant differences between only weed treatments, because of some taller weeds than soybean. At the middle level of canopy, all simple effects were significant in both years but interaction between nitrogen levels and varieties was significant in the first year of experiment. The least transmitted light was seen in urea treatments (Fig. 1). There were significant differences between two varieties for light interception (Fig. 2). Williams has wide leaflets with more surface than Harcor and its leaf angle is lesser than Harcor, so transmitted light to middle layer of its canopy was lesser than Harcor. Amounts of light in the middle of Williams' canopy were
 | |
| Fig. 2: | Variety effects on light interception at bottom and middle layer of canopy, left: first year and right: second year |
At the bottom level, Williams, canopy had only 6% of top intercepted radiation (Fig. 2). Effect of different N levels (Fig. 5) and weeds (Fig. 3) on the bottom level of canopy (soil surface) was very significant. In the first year interaction between nitrogen and variety was significant (Fig. 4), but in the second year interaction between nitrogen and weeds with interaction between variety and weeds were significant (Table 2).
| Table 2: | Mean of nitrogen* weed and variety* weed combinations for the PAR at the bottom layer of canopy in the second year of experiment (p<0.01) |
 | |
| Values with different superscripts letter(s) are significantly different at p<0.05 | |
 | |
| Fig. 3: | Weed effects on light interception at bottom and middle layer of canopy. Left: first year and right: second year |
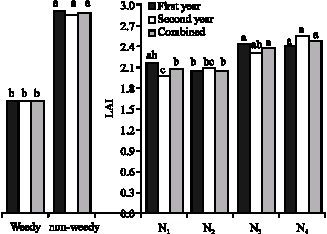
Leaf area index: Only simple effects of nitrogen and weeds had significant effects on LAI (Fig. 6). Urea because of its fast solubility in the soil was better than simbionts in leaf growth and expansion. Weed interference decreased soybean LAI in the first and second years about 44.4 and 43.5%, respectively. Van-Acker et al. (1993), Geddes et al. (1979) and Graham et al. (1988) have shown that soybean LAI decreases in weed competition.
LAI and light absorption increase in the canopy are related together by Light Interception Efficiency (LIE). LIE is defined as LI / LAI, (Board and Harville, 1992), which LI is light interception and calculated with this equation:
| LI = 1-[(I) / (I°)] | Purcell et al. (2002) |
| Where: |
| I | = | Quantity of radiation at the defined layer of canopy |
| I° | = | Radiation amount at the top surface of canopy |
A relation between LAI and LIE was found at the middle layer (Fig. 7). This is a negative relation, so by increasing LAI, LIE is declined. On the other hand, weed interference also increases total LAI (crop and weed LAI)
 | |
| Fig. 4: | Nitrogen and variety combination effects on light interception at bottom and middle layers of canopy (N is nitrogen level and V is variety) |
 | |
| Fig. 5: | Nitrogen effects on light interception at bottom and middle layers of canopy, Left: first year and right: second year (N is nitrogen level) |
and causes to faster decrease in LIE. Nitrogen applying in the weedy condition can strongly decrease crop LIE, but with the good control of weeds it decreases slowly.
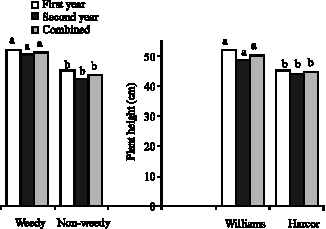
Root-shoot growth: Nitrogen sources had significant effect on plant height in the first year but no differences were seen in the second year. Combined analysis showed that effect of nitrogen on plant height depends upon climate changes in different years. Williams Was higher than Harcor and weed interference increased its height (Fig. 8). This can be resulted from plants competition for light (Board, 2000; Bullock et al., 1998; Bennett and
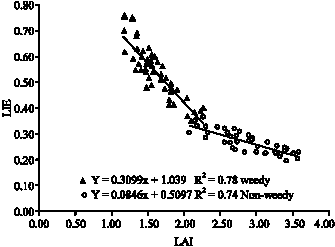 | |
| Fig. 7: | Relation between leaf area index and light interception efficiency in weed free and weed interference plots |
Shaw, 2000). Likely, changes in auxin and gibberellin concentration in plants due to light stress were the main reason for increasing plant height.
A relation between plant height and light interception in the middle strata of soybean,s canopy is shown in Fig. 9. Increasing height causes increase in light absorption by upper leaves, so bottom leaves senescence and die resulting end season LAI decline.
Root length only affected by different levels of nitrogen (Fig. 10). Singleton and Van Kessel (1987) introduced an interesting theory that called selectively partitioning of photosynthate in soybean. This theory can explain root growth in response to different nitrogen levels.
Fresh and dry weights of root nodules: Effect of weeds on root nodules was significant (Table 3). Weed competition decreased nodules weight (Fig. 11). There is little
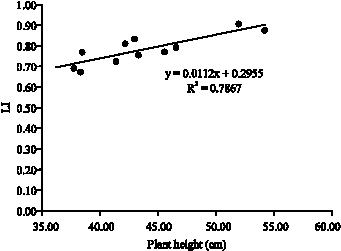 | |
| Fig. 9: | Relation between plant height and light interception at middle layer of mixture canopy (soybean and weeds) |
Stress conditions can change the amount and quality of nod gene stimulators that necessary for symbiosis and released from fabaceae plants root (Rengel, 2002). Activity levels of glutamine synthetase and glutamate synthase in varieties of fabaceae plants genetically are different that influence on rate and effectiveness of symbiosis (Caba et al., 1993). Dangaria et al. (1994) showed that pea varieties are different in number and weight of nodules. Israel (1981) found that two varieties of soybean significantly different in number of nodules, but their dry weight are constant. Pan et al. (2002) have reported
| Table 3: | Combined ANOVA for nodule weights |
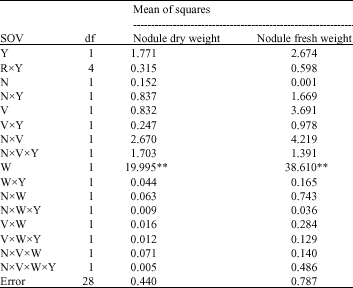 | |
| **: Indicates significant differences at 1% probability level, Y, R, N, V and W respectively refer to year, replication (block), nitrogen, variety and weed. In this table two levels of N were omitted because they had no bacteria treatment | |
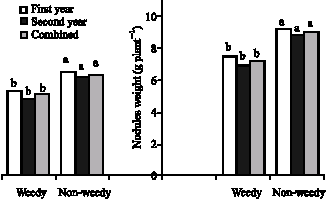 | |
| Fig. 11: | Weeds effects on nodules dry (left) and fresh (right) weights |
CONCLUSION
Although soybean genotypes are different in light interception efficiency, agronomical methods can significantly affect on their light absorption. One of the most important methods is weed control and rescue of crops from nutrient depletion. In addition, leaf area of weeds can absorb amounts of light and this cause a LIE decrease in mixed canopy. Weeds are hungry for elements specially nitrogen and use it very fast. Weed control is an economical way to farmers who want to avoid increasing fertilizer costs. This can promote root growth and its nodule activity to fix more nitrogen in comparison of weedy condition.
REFERENCES
- Bennett, C.A. and D.R. Shaw, 2000. Effect of Glycine max cultivar and weed control on weed seed characteristics. Weed Sci., 48: 431-435.
Direct Link - Board, J., 2000. Light interception efficiency and light quality affect yield compensation of soybean at low plant populations. Crop Sci., 40: 1285-1294.
Direct Link - Board, J.E. and B.G. Harville, 1992. Explanations for greater light interception in narrow vs. wide-row soybean. Crop Sci., 32: 198-202.
Direct Link - Bullock, D., S. Khan and A. Rayburn, 1998. Soybean yield response to narrow rows is largely due to enhanced early growth. Crop Sci., 38: 1011-1016.
Direct Link - Bussan, A.J., O.C. Burnside, J.H. Orf, E.A. Kistau and K.J. Puettmann, 1997. Field evaluation of soybean genotypes for weed competitiveness. Weed Sci., 45: 31-37.
Direct Link - Caba, J.M., C. Lluch and C. Ligero, 1993. Genotypic differences in nitrogen assimilation in Vicia faba: Effect of nitrate. Plant Soil, 151: 167-174.
CrossRef - Corre-Hellou, G., N. Brisson, M. Launay, J. Fustec and Y. Crozat, 2007. Effect of root depth penetration on soil nitrogen competitive interactions and dry matter production in pea-barley intercrops given different soil nitrogen supplies. Field Crops Res., 103: 76-85.
CrossRef - Ghosh, P.K., M. Mohanty, K.K. Bandyopadhyay, D.K. Painuli and A.K. Misra, 2006. Growth, competition, yields advantage and economics in soybean/pigeonpea intercropping system in semi-arid tropics of India: II. Effect of nutrient management. Field Crops Res., 96: 90-97.
CrossRef - Graham, D.L., J.L. Steiner and A.F. Wiese, 1988. Light absorption and competition in mix sorghum-pigweed communities. Agron. J., 80: 415-418.
Direct Link - Greci, I., P.H. Vivian, G. Hilbert, S. Milin, T. Robert and J.P. Gaudillere, 2007. Effect of light and nitrogen supply on internal C: N balance and control of root-to-shoot biomass allocation in grapevine. Environ. Exp. Bot., 59: 139-149.
CrossRefDirect Link - Jannink, J.L., J.H. Orf, N.R. Jordan and R.G. Shaw, 2000. Index selection for weed suppressive ability in soybean. Crop Sci., 40: 1087-1094.
Direct Link - Ofosu Budu, K.G., K. Fujita, T. Gamo and S. Akso, 1993. Dinitrogen fixation and nitrogen release from roots of soybean cultivar bragg and its mutants nts1116 and nts1007. Soil Sci. Plant Nutr., 39: 497-506.
Direct Link - Pan, B., J.K. Vessey and D.L. Smith, 2002. Response of field-grown soybean to co-inoculation with the plant growth promoting rhizobacteria Serratia proteamaculans or Serratia liquefaciens and Bradyrhizobium japonicum pre-incubated with genistein. Eur. J. Agron., 17: 143-153.
CrossRef - Purcell, L.C., R.A. Ball, J.D. Reaper and E.D. Vories, 2002. Radiation use efficiency and biomass production in soybean at different plant population densities. Crop Sci., 42: 172-177.
Direct Link - Rao, M.S.S., A.S. Bhagsari and A.I. Mohammed, 2002. Fresh green seed yield and seed nutritional traits of vegetable soybean genotypes. Crop Sci., 42: 1950-1958.
Direct Link - Graham, P.H., M. Hungria and B. Tlusty, 2004. Breeding for better nitrogen fixation in grain legumes: Where do the rhizobia fit in?. Crop Manage.,.
CrossRef - Sinclair, T.R. and C.T. de Wit, 1976. Analysis of the carbon and nitrogen limitations to soybean yield. Agron. J., 68: 319-324.
CrossRefDirect Link - Sinclair, T.R., L.C. Purcell, C.A. King, C.H. Sneller, P. Chen and V. Vadez, 2007. Drought tolerance and yield increase of soybean resulting from improved symbiotic N2 fixation. Field Crops Res., 101: 68-71.
CrossRef - Singleton, P.W. and C. Van-Kessel, 1987. Effect of localized nitrogen availability to soybean half-root systems on photosynthate partitioning to roots and nodules. Plant Physiol., 83: 552-556.
Direct Link - Song, L., B.J. Carroll, P.M. Gresshoff and D.F. Herridge, 1993. Field assessment of supernodulating genotypes of soybean for yield, N2 fixation and benefit to subsequent crops. Soil Biol. Biochem., 27: 563-569.
CrossRef