
M. Abdulla Al-Hashem
Department of Biological Sciences, Kuwait University, P.O. Box 5969, Safat, State of Kuwait
P.F. Brain
Department of Biological Sciences, Swansea University, Swansea, SA2 8PP, UK
S. Ahmad Omar
Food Resources Division, Kuwait Institute for Scientific Research, P.O. Box 13109, Safat, State of Kuwait
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 4 | Page No.: 589-594







ABSTRACT
An attempt was made to study the effects of oil pollution in a desert location (the Greater Al-Burgan oil fields, an area damaged in the second Gulf War) in Kuwait on the behaviour of the Sand lizard A. scutellatus. Polluted sites with apparently different degrees of contamination (namely tar mat, soot and clear sites) were compared with control areas outside this region. Between 2002 and 2003, ten lizards (5 of each sex) on each polluted and each control site were observed in the field at a time of the year when they were highly active. Air, substrate and burrow temperatures were recorded and lizards were monitored for their morning emergence times, as well as their basking and foraging activities. The present study confirmed that the morning emergence times and the basking behavior varied in sand lizards among the different pollution site categories. Physical changes in the tar mat sites caused the substrate temperatures in these locations to rise more quickly in the morning in response to solar gain than was the case in the other sites. This gives lizards in these locations the opportunity to emerge earlier and to start eating more quickly, giving them an energetic advantage (perhaps, in turn, influencing their rates of growth and fecundity). The clear sites had the next earliest emergence and were the next hottest but it is difficult to account for this in terms of the physical characteristics of this site. The basking times were clearly shorter on the dark soot and tar mat sites that appeared to have higher solar gain than control or clear sites. There did not appear to be any obvious differences in foraging activity of lizards in the different locations. It appears that some aspects of simple behaviour in these lizards provides a reliable, noninvasive indices for assessing oil pollution in desert locations. The precise impact of these changes in these reptiles on their long-term viability needs to be evaluated.
PDF Abstract XML References Citation
How to cite this article
M. Abdulla Al-Hashem, P.F. Brain and S. Ahmad Omar, 2008. Effects of Oil Pollution at Kuwait`s Greater Al-Burgan
Oil Field on the Timing of Morning Emergence, Basking and Foraging Behaviors
by the Sand Lizard Acanthodactylus scutellatus. Pakistan Journal of Biological Sciences, 11: 589-594.
DOI: 10.3923/pjbs.2008.589.594
URL: https://scialert.net/abstract/?doi=pjbs.2008.589.594
DOI: 10.3923/pjbs.2008.589.594
URL: https://scialert.net/abstract/?doi=pjbs.2008.589.594
INTRODUCTION
The Iraqi invasion of Kuwait on 2 August 1990 and the subsequent military activity left many scars on the desert ecosystem. Kuwait`s environmental problems were exacerbated by the burning of oil wells as the Iraqi troops withdrew. Over 730 oil wells were set on fire at this time. More than 80% of the wells burned while the rest gushed oil to the soil surface, covering wide areas of the desert (Omar et al., 2000). The oil fires released particles, organic and inorganic gases, Hydrocarbons (HCs) and oil droplets (Al-Hassan, 1992). Oil spills, aerosol deposits and seawater (used in attempts to extinguish the flames) all had adverse effects on the desert ecosystem. The explosion of the oil wells in Burgan and Ahmadi produced enormous volumes of soot and unburned oil in the form of oil-mist that was carried to distant locations over an area of approximately 49.15 km2 (Al-Ghunaim, 1997).
The resulting oil lakes became a severe, lasting source of pollution in this desert location. The lakes were formed by the discharge of oil from damaged wells that acted as gushers and burning wells, whose discharge rate was greater than could be consumed by the flames (such that the spray of oil finally landed back on the ground). The oil subsequently collected on the ground and ran into streams, following slopes and contours of desert topography. These running streams coalesced to form lakes of oil (Al-Hassan, 1992).
Lizards are important components of terrestrial ecosystems, forming an important link in food chains between invertebrate prey and predatory vertebrates such as birds and snakes (Lambert, 1997a, b). Lizards have rarely been used, however, as bioindicators of pollution for a variety of reasons, including the difficulty in sampling sufficient numbers and these reptiles relative lack of economic importance (Loumbourdis, 1997). Since, however, invertebrates are the prey of many lizards, the uptake of any chemical contaminants ingested by invertebrates is an important pathway by which pollutants enter the bodies of lizards. These chemicals can also result from the incidental ingestion of soil (small stones are often found in the digestive tracts of the lizards and are used to break up gut contents as in bird gizzards). Lizards have consequently been advocated as potential bioindicators of pesticides entering the environment (Lambert, 1993).
The Sand lizard A. scutellatus was chosen to assess some of the lasting impacts of oil pollution on desert fauna because it has a wide distribution. This species is a typical medium-sized lacertid with a cylindrical body and with well-developed limbs (Leviton et al., 1992; Salvador, 1982). The tail is long and the head is wide with an elongated pointed snout. A fringe of scales on the trailing edge of each (especially the 4th) toe, facilitates locomotion on loose sand and gives these animals their common name of fringe-toed lizards. This species is insectivorous, feeding mainly on ants, flies, small beetles and insect larvae (Perry and Dmiel, 1994).
It has been claimed that behavioural changes are amongst the most sensitive indicators of environmental contamination (Brain et al., 1994). Because no data is available on the effect of oil pollution on the behaviour of reptiles, a prime purpose of this initial study was to ascertain whether the behaviour of A. scutellatus reflected the level of oil pollution in desert locations.
The Greater Al-Burgan oil field has an area of 349.65 km2 and lies 20 km to the south of Kuwait City. Types of contaminated soils have been categorized on the basis of simple ground observations. The types identified have been designated as tar mat, soot and clear. The tar mat areas have a soil surface that is solidified by oil forming a crust about 1 cm thick that can be peeled off the underlying apparently clean soil. The soot areas have particulate black hydrocarbon deposits within the upper layer of soil to a depth of 1-8 mm. The clear sites have no visual evidence of soil pollution. Contamination may be continuous or discontinuous.
MATERIALS AND METHODS
Two sites for each category of contaminated soils were located in the oil field. Two comparable areas well outside the oil field were used as controls. The control area (Sulaibiya) is an agriculture Research Station at Kabd which was established in 1975. It is a fenced reserve protected from livestock and human interference by restricted access. Consequently, the area is highly vegetated, especially in Spring. The research station covers a total area of 20 km2, being 4 km (east to west) x 5 km (north to south).
Between 2002 and 2003, field observations were carried out on five lizards on 2 areas of each type of polluted site and 2 control sites (the sexes were balanced). Only mark-bearing lizards were used to overcome the problem of data repetition involving the same lizard (live specimens of sand lizards were permanently marked by toe clipping but easy visual recognition was facilitated by painting bands on the animals back using nail polish) (Al-Hashem, 2006). Individual lizards were watched, with 7x50 binoculars (Pentax, Asahi Optical Co., Ltd., Japan) where necessary, for extended periods of the year but mainly between February and April when these reptiles are maximally active. Three readings were taken for each observed lizard and average readings was recorded. The air, substrate and burrow temperatures were also recorded using mercury and thermocouple thermometers (Model No. 8528-10, Diqi-Sense, Cole-Palmer Instrument Company, Chicago, Illinois USA). These temperatures were measured in 2 replicates at each study site 3 times for each observed lizard and the average reading was recorded. The individual temperatures were taken 30 times at each site category over 30 days. These temperatures were taken during the times when the lizards were observed at each study site. Burrow temperature was taken by inserting the probe of the thermocouple thermometer to a depth of 8 cm into the lizards burrow.
Activities of A. scutellatus were monitored by visually searching the study areas each day at 08:00, 10:00, 13:00 h local time during the study period. When the weather became warm during February to April, monitoring was started one hour earlier. The study sites were slowly walked following an established route and each animal seen was watched for an extended period, so its behaviour could be quantified.
Each lizard was observed 3 times for morning emergence, basking and foraging activities. The average of these 3 measurements was recorded for each lizard, then the mean of the 10 averages (of 10 lizards) was taken for each study site (the sex of the lizard did not appear to influence this variable). This procedure was also performed with air, substrate and burrow temperatures. During each observation, these individual temperatures were measured, (again 3 readings for each lizard with the average for these 3 readings being recorded). The mean of 10 averages (of 10 lizards) was taken for each study site for each individual temperature.
The data were analyzed with Minitab, version 13.32 (2003) and SPSS, version 11.0 (2001). Data analysis was performed using the parametric one-way analysis of variance (ANOVA) test. Parametric post hoc Scheffe' tests were used to compare pairs of study sites.
RESULTS AND DISCUSSION
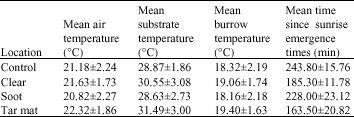
At the time of the study, emergence of A. scutellatus was observed when air temperatures were around 20-25°C and substrate temperatures 25-35°C. Emergence could take place earlier or later depending on the study site and on the weather (degree of cloud cover and/or wind conditions), but most emergence occurred 2 to 3 h after sunrise in the tar mat and clear sites but about an hour later in the other study sites. In most cases, the lizard would first extend its head, then, after few minutes, it would expose the entire body (Table 1).
Although there was some variability, there was no significant overall difference on ANOVA in air temperature between the study sites, which means that air temperature could not account for any emergence time variation. There was also no significant overall variance of substrate temperatures that were taken during morning emergence between the study sites. One-way ANOVA also showed no significant difference in burrow temperatures between the study sites. One-way ANOVA revealed, however, significant differences in the emergence times seen at the different sites (F3,36 = 40.78, p<0.0001).
The post hoc Scheffe' tests showed no difference in the minutes since sunrise emergence times of sand lizards between the control and the soot sites. The lizards from the control and the soot sites emerged, however, significantly later than counterparts at the clear and the tar mat sites (p<0.0001). This suggests that animals at the last two sites can start feeding earlier. The view that clear sites are less polluted than the soot sites and that the tar mat sites are the most polluted has been recently shown by Al-Hashem et al. (2007) to be a gross over-simplification. All the polluted sites revealed similar levels of HCs in soil and lizard tissue samples and all produced much higher levels recorded in samples from the control sites.
Following the morning emergence, A. scutellatus would bask in the sunlight to raise its body temperature to the level required for the normal activity. Basking usually took place in close proximity to the lizards burrow. Lizards generally lie motionless close to their burrows but were alert and would retreat if disturbed. In order to warm up, the lizard widely spreads its ribs and orients itself so the long axis of its body is perpendicular to the rays of the sun with the legs stretched out. In this situation, the sun
| Table 1: | Mean±SD air, substrate and burrow temperatures as well as morning emergence times (N = 10) at the different study sites |
 | |
| Table 2: | Mean±SD air and substrate temperatures as well as basking durations (N = 10) at the different study sites |
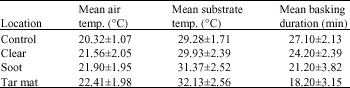 | |
strikes the lizards entire dorsal surface and the reptile intercepts the maximum amount of solar radiation and casts a relatively large shadow on the ground. In contrast, when the lizard is hot and trying to minimize the amount of solar radiation received, it compresses its ribs against its body and faces directly into the sun. In this position the sun strikes perpendicularly only on the lizards head and shoulders and the reptile casts a much smaller shadow.
The amount of time allocated to basking was measured for thirty minutes for each lizard after morning emergence at the types of oil polluted and the control sites (Table 2).
One-way ANOVA was used to investigate the basking seen on the varied sites. Basking duration showed significant variation between the study sites (F3,36 = 16.87, p<0.0001).
The post hoc Scheffe' tests showed that there was no difference in basking period of sand lizards at the control and the clear sites. There was a significant difference between the control and the soot sites (p<0.001), the control and the tar mat sites (p<0.0001) and the clear and the tar mat sites (p<0.001) in basking period of sand lizards. In essence, the mean basking duration was shortest at the tar mat sites and it was shorter in the soot sites than in the clear sites. Mean basking duration was longest at the control sites.
There was no significant variation of air temperatures between the study sites, indicating that variations in air temperatures did not account for the varied basking durations. The substrate temperatures were measured by mercury and thermocouple thermometers when air temperatures were recorded after morning emergence at the different study sites. The means showed a significant variation (F3,36 = 3.15, p<0.03). Consequently, variations in substrate temperatures could be an important factor in producing this difference among the study sites (control sites and the tar mat sites are both shaded by vegetation but the tar mat sites appeared to absorb solar radiation more effectively). They may function a little like the polytunnels used in agriculture.
As used by Pianka et al. (1979), the mean number of Moves per Minute (MPM) made were used to compare the foraging strategies of A. scutellatus at the different pollution sites. MPM in this study was determined after the animal basks in the sun and becomes active, any movement included to capture flying insect or creeping insect larvae or walking insects was recorded. During observations, care was taken to avoid disturbing the focal animal or of observing the same animal twice. Consequently, only 10 marked lizards from each study site were included in these measurements. Although it was impossible to completely exclude all other activities during such a study, data collected while animals appeared involved in other activities such as fighting, mating, thermoregulation and predation avoidance were eliminated from the analysis. The mean MPMs at the 4 different sites are shown in Table 3.
The one-way ANOVA test revealed no significant variance in foraging activity at the different study sites (F3,36 = 0.77, p = 0.52). It worth noting, however, that the lizards at the tar mat site showed the most MPM followed by the lizards from the control site. The lizards from the soot site showed the lowest MPM value.
This study shows that A. scutellatus populations at the clearly oil polluted and the non polluted (control) sites show differences in their daily emergence and basking behaviour. In the tar mat sites, the lizards emerged earlier than from any of the other sites. In spite of the ambient temperature conditions being similar, field observations provided an indication that substrate temperature varied among the different sites of study and had an impact on the emergence and basking of A. scutellatus. The tar mat sites exhibited the highest substrate temperature. It seems likely that the physical characteristics of the site (black and with a crust) causes such areas to warm quickly perhaps changing the behaviour of associated reptiles. It is more difficult to account for the early emergence times of lizards from the clear sites (these were neither black nor crusted). They did, however, also have relatively high substrate and burrow temperatures.
Many researchers Pianka (1966),Perry and Pianka (1997) and McBrayer and Reilly (2002) have recognized two basic modes of foraging used by carnivorous lizards, commonly called Sit-and-Wait (SW) and Widely Foraging (WF) styles. SW foragers tend to capture mobile prey whereas WF foragers generally feed on sedentary prey (Huey and Pianka, 1981; Huey et al., 2001). The frequency
| Table 3: | Mean±SD MPMs (N = 10) at the different study sites |
 | |
of prey that wander by a SW predator is positively correlated with prey density, so the success of a SW forager declines with prey scarcity (Schoener, 1969;Cooper, 1995). SW foraging is more energetically conservative than active foraging (Huey et al., 1983; Cooper, 2000) and is generally favoured in reptiles when mobile prey are sufficiently abundant. When mobile prey are limited or prey abundance is reduced, active searching appears to be favoured. The foraging mode varies between species of reptiles and is correlated with patterns of habitat and prey use (Huey and Pianka, 1981; Petranka, 1998). The foraging movements of WF reptiles are naturally more extensive than those of SW counterparts (Pietruszka, 1986). There have been few studies on the foraging behaviour of lizards in the wild, so there is an incomplete understanding of the detailed behaviours of WF and SW predators (Day et al., 1999). Arnold (1984), Theodoratus and Chiszar (2000) and Clark (2004) explained that vision is a main sensory system needed for SW predators while hearing and olfaction are often involved in WF predators. Vulnerability to visual predators is low in SW foragers because lizards are often immobile and can forage without detection in exposed positions if they possess good crypsis. Vulnerability to visual predators is, however, higher in WF strategists because their movement draws attention to these lizards. Consequently, unless predators are absent, active searching is only likely to take place in protected situations such as in vegetative cover. Prey mobility must be high if these are to reach SW foragers and their visibility should be high as they are often detected at considerable distances (Arnold, 1984; Theodoratus and Chiszar, 2000; Clark, 2004).
Although the lizards at the tar mat site showed the greatest number of MPM in foraging at these sites, the result was not significantly different from that for lizards at the other sites.Perry et al. (1990) measured the MPM for A. scutellatus, finding a mean of 1.01. In this present study, the mean of MPM was 0.199 for A. scutellatus from the tar mat sites and 0.172 for the controls. The difference between these results in the two studies might be related to food resource availability or the biodiversity of prey species. Various diets will be assorted with different efficiencies and energy requirements. The predators may increase their MPM to fulfill their dietary requirements or may reduce this index because the food is abundant and needs little energy to be consumed. A proximate reason for a predator to limit its movements could be reducing its own chance of being predated. Risk of predation usually increases with the amount of movement so the SW strategy of lizards avoids moving in order to not draw the attention of their own predators. Huey and Pianka (1981) suggested that SW strategists generally rely on crypsis to avoid being detected. The MPM study confirms that the lizard persists as a SW at all the locations and does not apparently have to increase its active foraging.
It appears that the morning emergence time and basking activity of A. scutellatus provides, at least in some locations, a sensitive yet non-destructive (in terms of fauna) indicator of oil pollution. In general, obvious pollution resulted in the lizards being active earlier and potentially for longer. The foraging activity appears to be a more fixed element in the repertoire of these reptiles (perhaps reflecting the hunting style). It seems important to assess just what impact any behavioural changes and the exposure to pollutants have on the long-term viability of these animals.
REFERENCES
- Al-Hashem, M., P.F. Brain and S. Omar, 2007. Effects of oil pollution at Kuwait's Greater Al-Burgan Oil Field on polycyclic aromatic hydrocarbon concentrations in the tissues of the desert lizard Acanthodactylus scutellatus and their ant prey. Ecotoxicology, 16: 551-555.
Direct Link - Clark, R.W., 2004. Timber Rattlesnakes (Crotalus horridus) use chemical cues to select ambush sites. J. Chem. Ecol., 30: 607-617.
Direct Link - Cooper, W.E., 2000. An adaptive difference in relationship between foraging mode and responses to prey chemicals in two congeneric scincid lizards. Ethology, 106: 193-206.
Direct Link - Huey, R.B., E.R. Pianka and L.J. Vitt, 2001. How often do lizards run on empty? Ecology, 82: 1-7.
Direct Link - Lambert, M.R.K., 1997. Environmental effects of heavy spillage from a destroyed pesticide store near Hargeisa (Somaliland) assessed during the dry season, using reptiles and amphibians as bioindicators. Arch. Environ. Contam. Toxicol., 32: 80-93.
CrossRefDirect Link - Loumbourdis, N.S., 1997. Heavy metal contamination in a lizard, Agama stellio, compared in urban, high altitude and agricultural, low altitude areas of North Greece. Bull. Environ. Contam. Toxicol., 58: 945-952.
Direct Link - Perry, G. and R. D'miel, 1994. Reproductive and population biology of the fringe-toed lizard, Acanthodactylus scutellatus, in Israel. J. Arid Environ., 27: 257-263.
CrossRefDirect Link - Schoener, T.W., 1969. Models of optimal size for solitary predators. Am. Nat., 103: 277-313.
Direct Link - Theodoratus, D.H. and D. Chiszar, 2000. Habitat selection and prey odor in the foraging behavior of Western rattlesnakes (Crotalus viridis). Behaviour, 137: 119-135.
Direct Link