
R. Peyghan
Faculty of Veterinary, Shahid Chamran University of Ahvaz, P.O. Box 145, Ahvaz, Iran
M.D. Powell
School of Aquaculture, Tasmanian Aquaculture and FisheriesInstitute, The Cooperative Research Centre for Sustainable Development of Fin Fish Aquaculture, University of Tasmania, Locked Bag 1370, Launceston, Tasmania, 7250, Australia
M.R. Zadkarami
Department of Statistics, Faculty of Mathematics,Shahid Chamran University, Ahvaz, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 41-47







ABSTRACT
Neoparamoeba pemaquidensis believed to be the most prevalent parasite of Atlantic salmon industry in Australia. In the present study, the in vitro effects of crude extract of garlic and metronidazole were investigated using a primary culture toxicity assay. Garlic extract appeared to be completely effective at killing a cultured strain (NP251002) of Neoparamoeba pemaquidensis in vitro at a dilution of 1:100 with in 24 h. The number of viable Amoebae after using garlic extract in lower dilutions (1:200, 1:400, 1:800, 1:1000) for 24 h, also were significantly lower than in the control group. Garlic extract was also efficacious at killing wild type Amoebae that isolated from the diseased fish showing clinical signs of AGD. Metronidazole had no clear effect against Neoparamoeba pemaquidensis (NP251002) even in a concentration of 50 mg L-1 for 24 h. However some morphological changes have occurred in metronidazole-treated Amoebae after 5 days of exposure.
PDF Abstract XML References Citation
How to cite this article
R. Peyghan, M.D. Powell and M.R. Zadkarami, 2008. In vitro Effect of Garlic Extract and Metronidazole Against Neoparamoeba pemaquidensis, Page 1987 and Isolated Amoebae from Atlantic Salmon. Pakistan Journal of Biological Sciences, 11: 41-47.
DOI: 10.3923/pjbs.2008.41.47
URL: https://scialert.net/abstract/?doi=pjbs.2008.41.47
DOI: 10.3923/pjbs.2008.41.47
URL: https://scialert.net/abstract/?doi=pjbs.2008.41.47
INTRODUCTION
Amoebic Gill Disease (AGD) is an important disease of Atlantic salmon (Salmo salar) in Tasmania and has been reported from other areas (Munday et al., 2001). The morphology of Neoparamoeba pemaquidensis, the causative agent of AGD, has been extensively studied and a new species N. branchiophila has also been identified in AGD infections (Dykova et al., 2005). Typical pathological changes of fish infected with Neoparamoeba pemaquidensis are the hypertrophy and desquamation of surface epithelial cells within the immediate vicinity of attachment, hyperplasia and thickening of secondary lamellae as well as edema of the epithelium. The fishes are highly susceptible to infection in the sea water and the resulting gill damage and its influence on respiratory and acid-base balance contributes considerably to the lethal effect caused by the parasite (Powell et al., 2000; Powell and Nowak, 2003; Adams and Nowak, 2003).
There are several reports about the treatment of AGD affected fishes (Zilberg et al., 2000; Parsons et al., 2001; Clark et al., 2003; Roberts and Powell, 2004; Harris et al., 2004) but, the only effective commercial scale treatment for AGD to date is freshwater bathing that can be effective in the removal of majority of Neoparamoeba sp. from the gills of the affected fishes (Parsons et al., 2001). However, there are some reports about the effectiveness of other compounds as either bath treatments or as potential in feed amoebocides (Powell et al., 2003; Powell and Clark, 2003; Powell et al., 2005). Freshwater bathing to successfully avoid AGD is a critically important industry concern in term of production costs. Therefore the development of more effective drugs to control AGD in cultured fish is being sought.
Garlic (Allium sativum) is known as a potent medicine with broad therapeutic properties ranging from antibacterial to anticancer, antiparasitic and anticoagulant (Harris et al., 2001a; Ankri and Mirelman, 2001). Crude extracts of garlic and many other plants have been used as chemotherapeutics in different investigations (Emeruwa, 1982; Tona et al., 1998; Chinoy et al., 1995; Udoh and Kehinde, 1999; Asres et al., 2001; Buchmann et al., 2003; Ijah and Oyebanji, 2003; Lawson, 1996). The freshly cut slices of garlic bulb have sulfur-containing compounds such as allicin, the primary contributor of the pungency and medicinal properties of garlic (Ankri and Mirelman, 2001). Whole garlic typically contains about 1% allein, which is the odorless precursor of allicin. Allinase, which is an enzyme that is located within a separate compartment in natural garlic, converts allein into the biologically active allicin following the crushing of garlic and the contact between the enzyme and the substrate (Langa et al., 2004). Purification of allicin is a difficult tusk and was done by differential adsorption of the reaction components on a neutral polystyrene resin (Miron et al., 2004).
Other important sulphur-containing compounds present in garlic homogenate are allyl methyl thiosulphonate, 1-propenyl allyl thiosulphonate and L-glutamyl-S-alkyl-L-cysteine. Garlic oil consists of the diallyl (57%), allyl methyl (37%) and dimethyl (6%) mono to hexa sulphides. A typical commercial preparation of garlic oil contains 26% diallyl disulphide (DADS), 19% diallyl trisulphide (DATS), 15% allyl methyl trisulphide, 13% allyl methyl disulphide, 8% diallyl tetrasulphide, 6% allyl methyl tetrasulphide, 3% dimethyl trisulphide, 4% penta sulphide and 1% hexa sulphide. All polysulphides show antioxidant effects in vitro but this has yet to be confirmed in vivo. Diallyl polysulphide (S = 3 to 7) inhibits the formation of thiobarbituric acid reactive substances in vitro (Horie et al., 1992). Quantitative tests revealed that there were 17.8 n moles sulfhydryl (SH)/ml of 40 KD garlic protein localized in the parenchyma sheath cells and the cortical cells of garlic bulbs (Wen et al., 2004).
Metronidazole is the only 5-nitroimidazole approved for used in the United States and it is the drug of choice for treating giardiasis and amoebiasis (mainly referring to disease caused by Entamoeba histolytica) in many countries (Campanati and Monteiro-Leal, 2000). Both giardiasis and amoebiasis causative agents are anaerobic endoparasites and act differently from Neoparamoeba sp. which is an aerobic ectoparasite, but this parasite might also respond to metronidazole treatment.
In this study, the anti-amoebic effect of garlic extract as a natural extract and metronidazole as a potentially amoebostatic and anti-protozoal drugs were examined for their efficacy and toxicity to Neoparamoeba sp. in vitro.
MATERIALS AND METHODS
This study used Neoparamoeba pemaquidensis (NP251002) and amoeba isolated from Atlantic salmon, originated from one of Tasmanian hatcheries which were introduced at the Aqua center of school of Aquaculture, University of Tasmania, Launceston-Australia. The laboratory examinations and toxicity tests were done from October 2004 to January 2005.
Garlic extract preparation: Raw garlic homogenate prepared by crushing the 10 g garlic bulb in 100 mL water (water temperature: 20-22°C) in a pestle and mortar and passing the homogenate through paper filter and antibacterial paper (0.2 μm filter). For preparation of heated garlic extract, after the filtration of the extract, it placed in 80°C oven for 2 min. As a result of this heating, the transparent garlic extract changed to cloudy white. Forty microliter per milliliter of an antibiotic mixture: (Streptomycin 0.1 g, penicillin 0.1 g, carbenicillin 0.1 g, ampicillin 0.25 g, distilled water 10 mL) (Sigma, St. Louis, MO, USA) was added to the garlic extract and the control groups wells.
In vitro toxicity test with Neoparamoeba pemaquidensis (NP251002) Neoparamoeba pemaquidensis preparation: Neoparamoeba pemaquidensis (strain NP251002) were cultured on seawater malt yeast extract (Oxoid, Victoria, Australia) agar plates at 18°C. Amoebae were not fed additional bacterial cultures during the culture period.
Amoebae were harvested from the agar plate by washing and suspending them in 0.2 μm filtered seawater. The harvested Amoebae were counted using a haemocytometer (Neubauer, BS 748) and their viability assessed using trypan blue exclusion staining (Sigma) according Powell et al. (2003). The trypan blue staining was used for assaying the live Amoebae (live Amoebae do not taking up the trypan blue stain).
Fresh garlic extract toxicity test: Two hundred microlitres of cultured Amoebae solution was added to each well in a 96 well micro plate (Sarstedt-Australia). Twenty microlitres of garlic extract was added to each experimental group to make a final dilution of 1:100. By adding water, the serial dilution of 1:200, 1:400, 1:800, 1:1000, 1:1200 and 1:1400 were prepared and finally all of the amoeba solutions were incubated at 18.5°C for 24 h.
In other experiment, 200 μL of cultured Amoebae solution was added to each well in a 96 well micro plate (Sarstedt-Australia). Twenty microlitres of garlic extract was added to each experimental group to make a final dilution of 1:100. Dilution was incubated at 18.5°C for 8 h and amoeba number of each well were counted at 2 h intervals.
Heated garlic extract toxicity test: For preparation of heated garlic extract, after the filtration of the extract, it placed in 80°C oven for 2 min 200 μL of cultured Amoebae solution was added to each well in a 96 well micro plate (Sarstedt-Australia). Twenty microliter of the extract was added to each experimental group to make a final dilution of 1:100. By adding water, the serial dilution of 1:200, 1:400, 1:800 and 1:1000 were prepared and finally all of the amoeba solutions were incubated at 18.5°C for 24 h.
In vitro toxicity test with wild-type Amoebae Wild type Amoebae isolation: In order to confirm the efficacy of garlic extract on wild type of amoebae, Amoebae isolation from AGD-affected Atlantic salmon from an ongoing laboratory infection at University of Tasmania was conducted. Amoeba isolation from diseased fish was carried out according to Morrison et al. (2004).
Fresh garlic extract toxicity test: Two hundred microlitres of cultured Amoebae solution was added to each well in a 96 well micro plate (Sarstedt-Australia). Twenty microlitres of garlic extract was added to each experimental group to make a final dilution of 1:100. By adding water, the serial dilution of 1:200, 1:400, 1:800 and 1:1000 were prepared and finally all of the Aamoeba solutions were incubated at 18.5°C for 24 h.
In other experiment, 200 μL of isolated Amoebae solution was added to each well in a 96 well micro plate (Sarstedt-Australia). Twenty microlitres of garlic extract was added to each experimental group to make a final dilution of 1:100. Dilution was incubated at 18.5°C for 8 h and amoeba number of each well were counted at 2 h intervals.
Metronidazole toxicity test: Using same method, 200 μL of amoeba suspension was continuously exposed to metronidazole (Sigma, St. Louis, MO, USA) in different concentrations (0.1-100 mg L-1) over 6 days. The number of Amoebae in all experimental and control groups was determined using a haemocytometer (Neubauer), after staining with 0.5% trypan blue-seawater mixture at a dilution of 1:5 (Powell et al., 2003). Six replicate counts were made with counting of nine large haemocytometer squares per replicate.
Light microscopic observation of amoebae: One hundred microlitres of amoeba suspensions from garlic exposed Amoebae (from 1:100 dilution) and metronidazole (from concentration of 100 mg L-1) exposed Amoebae was placed on to a glass slide for 1 h for adherence of amoebae. Non-adherence cells were washed off. The slides were fixed by 2 drops of methanol. After drying in room temperature, the slides were stained by 1:10 dilution of giemsa and methylene blue stain for 15 min. A Leica DC300F digital camera (Leica-Germany) mounted to a light microscope (Olympus) was used for image capture.
Statistical analysis: The data was transformed to the Log-units to see the real effect and was analyzed by one-way (data by groups) (ANOVA). Post hoc Tukey`s, alpha (0.05) test was used for mean comparisons where the difference was significant between the groups. Statistics for each analysis were based on cases with no missing data for any variable in the analysis. SPSS (Version 10.0, SPSS Science) were used for data analysis and presentation. Data are presented as mean±standard error of mean (SE).
RESULTS
Fresh garlic extract toxicity test:
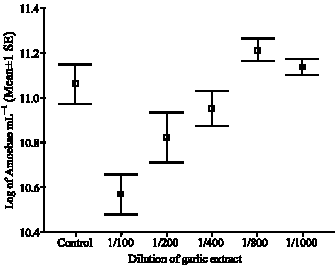
A: Neoparamoeba, NP251002: In the 24 h toxicity test, the Amoebae count for 1:100 and 1:200 dilutions was zero and in the 1:400, 1:800 and 1:1000 dilution of garlic solution was significantly lower then the control. At higher dilutions (1:1200 and 1:14000) there were no significant differences between garlic-exposed groups and the control (Fig. 1).
Numbers of live Amoebae (Neoparamoeba, Np251002) in the experimental group treated with garlic extract (dilution of 1:100) began to decrease significantly after 2 h of exposure and reached to zero after 8 h (Fig. 2).
Wild type Amoeba: The wild type Amoebae number count in the experimental group treated with 1:100 garlic extract also decreased significantly (Fig. 3). However, in other
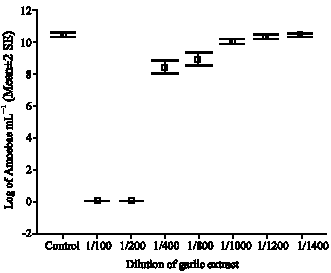 | |
| Fig 1: | In vitro toxicity test of Neoparamoba pemaquidensis (cultured strain NP251002) to various dilutions of fresh garlic extract in 24 h |
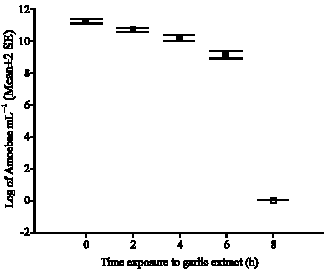 | |
| Fig 2: | In vitro toxicity test of Neoparamoba pemaquidensis (cultured strain NP251002) exposed to 1:100 dilution of fresh garlic extract in 8 h |
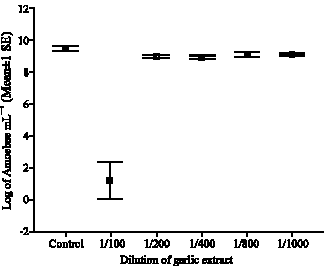 | |
| Fig 3: | In vitro toxicity test of wild type amoeba to various dilutions of fresh garlic extract in 24 h |
dilutions (1:400, 1:800 and 1:1000) there was not any significant difference between the experimental groups and the control. Numbers of live Amoebae (wild type) in the experimental group treated with garlic extract (dilution of 1:100) began to decrease significantly after 6 h of exposure (Fig. 4).
Heated garlic extract toxicity test:
In the toxicity test with heated garlic extract, the Amoebae count (Neoparamoeba, NP251002) in the control group significantly higher than the 1:100 dilution after 24 h exposure to heated garlic extract. However, at higher dilutions (1:400, 1:800 and 1:1000) garlic had not any significant effect in decreasing Amoebae count in 24 h (Fig. 5).
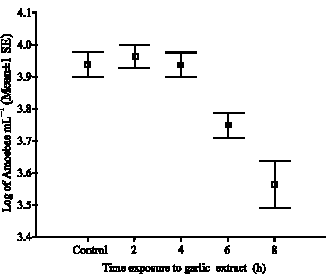 | |
| Fig 4: | In vitro toxicity test of wild type Amoeba exposed to 1:100 dilution of fresh garlic extract in 8 h |
 | |
| Fig 5: | In vitro toxicity test of Neoparamoba pemaquidensis (cultured strain NP251002) exposed to 1:100 dilution of heated garlic extract over 24 h |
Metronidazole toxicity test: Metronidazole had no significant effect against Neoparamoeba in a concentration of lower than 50 mg L-1 in 24 h. The Amoebae count, however, in the control group was significantly higher than the 100 mg L-1 group after 6 days of exposure (Fig. 6).
Morphological changes: Garlic extract caused cell vacuolization and swelling that lead to the rapture and death of the amoebae. These changes were not seen in Amoebae of the control groups. In comparison with control group amoebae, some morphological changes
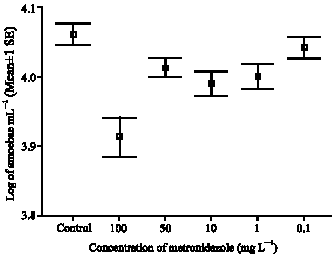 | |
| Fig 6: | In vitro toxicity test of Neoparamoba pemaquidensis (cultured strain NP251002) exposed to metronidazole in different concentrations in 6 days |
(abnormality of cell wall and the cytoplasm) in treated Amoebae were observed after 5 days exposure to metronidazole.
DISCUSSION
In this study garlic extract had an obvious antiamoebic effect. However, an effective treatment using natural sources would be less expensive then some other synthetic drugs. Moreover, this kind of product may be safe for fish and have less impact on the environment. Garlic (Allium sativum) has played an important medicinal role for centuries. Raw garlic homogenate is the major preparation of garlic that has been subjected to intensive scientific study. Allicin is the major thiosulphinate compound found in garlic homogenate (Benerjee et al., 2003). It is the one of the active principles of freshly crushed garlic homogenates that was found to exhibit antibacterial and antiparasitic activity by means of chemical reaction with thiol groups of various enzymes (Ankri and Mirelman, 2001). Inhibito-lethal activity against Giardia intestinalis was noted with crude extract of garlic at 25 μg mL-1 and lethal dosage was established as 50 μg mL-1 (Harris et al., 2001b).
In this study metronidazole had not any significant effect on Neoparamoeba sp. in concentration lower then 50 mg L-1. Metronidazole is used to treat a broad range of infections caused by anaerobic protists and bacteria. This drug is also effective against human amoebiasis. The selective cytotoxicity of metronidazole relies on biochemical properties of anaerobic organisms that are lacking in the aerobic cells. For aerobic cells the inhibitory concentration of metronidazole is two to three orders of magnitude higher than that for anaerobes (Land and Johnson, 1997). Some ultra structural studies are needed for further determination of morphological changes (abnormality of cell wall and the cytoplasm) of the Amoebae that were observed after treatment. Morphological changes of metronidazole may be the effect of DNA changes in the treated amoebae. Metronidazole appeared to have a role in inducing DNA damage and DNA breakage due to treatment in human lymphocytes through the futile cycle (Menendez et al., 2001; Re et al., 1997). However because the morphological changes in our study were not characterized by more specific staining and or ultra structural studies, further studies in this respect is recommended.
Garlic and metronidazole have not previously been investigated against N. pemaquidensis. This study showed that garlic extract strongly influenced the survival of the gill Amoebae in vitro. In the present investigation, it is showed clearly that the concentration of 100 mg L-1 is specifically important for effecting survival of N. pemaquidensis in 1 h. Although N. pemaquidensis can withstand relatively metronidazole, it is showed that higher concentration of metronidazole (100 mg L-1) kills the majority of Neoparamoebae causing AGD in salmon in vitro. Previous research has suggested that only fresh water can kill N. pemaquidensis (Clark, 2002). There are a lot of risk factors within the Tasmanian salmon aquaculture industry when fresh water are using for treatment. Bath duration typically ranges from 2 to 4 h, the water used to bathe fish is often hard and not all of the Amoebae are removed from the gills. Survival of some Amoebae on the gills during bathing either within the structure of the gill (Parsons et al., 2001) or in the gill mucus (Clark, 2002) would mean that freshwater resistant Amoebae may be developed. Other chemical treatments were variable in their ability to reduce the relative survival of gill Amoebae including N. pemaquidensis in vitro. Chloramine-T and hydrogen peroxide showed the most promise. Chlorine-based disinfectants such as chloramine-T primarily act by the release of hypochlorite (Booth and McDonald, 1988). The effects of these disinfectants on the Amoebae may be mitigated by the presence of mucus. This may be particularly important when using disinfectants to remove and kill Amoebae on the gills of fish. In the latter case, hypochlorite results in irritation and alteration in gill epithelial membrane permeability (Powell and Perry, 1998) resulting in a hyper secretion of bronchial mucus, potentially aiding the removal of the parasites.
In conclusion, we have demonstrated that, fresh garlic extract in dilution of 1:100 (concentration of about 10 g L-1) in vitro, is efficacious in killing the Neoparamoebae sp. after 8 h exposure in vitro. Therefore garlic extract or its active antimicrobial agent (Allicin) may be used alone or in combination with other effective drugs (or fresh water bath) in the treatment of amoebic gill disease in farmed Atlantic salmon. However, before that it is necessary to study the toxicity and pathological effect of garlic extract on Atlantic salmon and establish the efficacy of garlic extract for the removal of Neoparamoeba sp. from the gills of infected fish.
ACKNOWLEDGMENTS
We would like to thanks the contributions of Dr. P. Crosbie for supplying culture Amoebae (NP251002), some isolated amoeba and the antibiotics mixture. This work formed part of a project of Aquafin CRC and received funds from the Australian Government`s CRCs Program, the Fisheries R and D Corporation and other CRC Participants and was done as part of sabbatical study of first author funded by Shahid Chamran University, Ahvaz- Iran.
REFERENCES
- Adams, M.B. and B.F. Nowak, 2003. Amoebic gill disease: Sequential pathology in cultured Atlantic salmon, Salmo salar L. J. Fish Dis., 26: 601-614.
Direct Link - Ankri, S. and D. Mirelman, 1999. Antimicrobial properties of allicin from garlic. Microbes Infect., 1: 125-129.
CrossRefDirect Link - Asres, K., F. Bucar, E. Knauder, V. Yardley, H. Kendrick and S.L. Croft, 2001. In vitro antiprotozoal activity of extract and compounds from the stem bark of Combretum molle. Phytother. Res., 15: 613-617.
Direct Link - Benerjee, S.K., P.K. Mukherjee and S.K. Maulik, 2003. Garlic as an antioxidant: The good, the bad and the ugly. Phytother. Res., 17: 97-106.
CrossRefPubMedDirect Link - Buchmann, K., P.B. Jensen and K.D. Kruse, 2003. Effects of sodium percarbonate and garlic extract on Ichthyophthirius multifiliis theronts and tomocysts: In vitro experiments. North Am. J. Aquacul., 65: 21-24.
Direct Link - Chinoy, N.J., T. Dilip and J. Harsha, 1995. Effect of Carica papaya seed extract on female rat ovaries and uteri. Phytother. Res., 9: 169-175.
CrossRefDirect Link - Campanati, L. and L.H. Monteiro-Leal, 2000. The effect of the antiprotozoal drugs metronidazole and furazolidone on trophozoites of Giardia lamblia (P1 strai). Parasitol. Res., 88: 80-85.
Direct Link - Clark, G., M. Powell and B. Nowak, 2003. Effects of commercial freshwater bathing on re-infection of Atlantic salmon, Salmo salar with amoebic gill disease. Aquaculture, 219: 135-142.
Direct Link - Dykova, I., B.F. Nowak, P.B.B. Crosbie, I. Fiala and H. Peckova et al., 2005. Neoparamoeba branchiphila n. sp. and related species of the genus Neoparamoeba: Morphological and molecular characterization of selected strains. J. Fish Dis., 28: 49-64.
Direct Link - Emeruwa, A.C., 1982. Antibacterial substance from Carica papaya fruit extract. J. Nat. Prod., 45: 123-127.
CrossRefDirect Link - Harris, J.C., S.L. Cottrell and D.L. Plummer, 2001. Antigiardial drugs. Applied Microbiol. Biotechnol., 57: 614-619.
CrossRefDirect Link - Harris, J.C., S.L. Cottrell, S. Plummer and D. Lloyd, 2001. Antimicrobial properties of Allium sativum (garlic). Applied Microbiol. Biotechnol., 57: 282-286.
CrossRefDirect Link - Harris, J.O., M.D. Powell, M. Attard and T.J. Green, 2004. Efficacy of chloramine-T as a treatment for a treatment for amoebic gill disease (AGD) in marine Atlantic salmon (Salmo salar L.). Aquacul. Res., 35: 1448-1456.
Direct Link - Horie, T., S. Awazu, Y. Stakura and T. Fuwa, 1992. Identified diallyl poly sulfides from an aged garlic extract which protects the membrane from lipid per oxidation. Planta Med., 58: 468-469.
Direct Link - Ijah, U.J.J. and F.O. Oyebanji, 2003. Effects of tannins and polyphenols of some medical plants on bacterial agents of urinary tract infections. Global J. Pure Applied Sci., 9: 193-198.
Direct Link - Land, K.M. and P.J. Johnson, 1997. Molecular mechanisms underlying metronidazole resistance in trichomonads. Exp. Parasitol., 87: 305-308.
CrossRef - Langa, A., M. Lahava, E. Sakhninia, I. Barshack and H.H. Fidder et al., 2004. Allicin inhibits spontaneous and TNF-a induced secretion of proinflammatory cytokines and chemokines from intestinal epithelial cells. Clin. Nutr., 23: 1199-1208.
CrossRef - Menendez, D., E. Rojas, L.A. Herrera, M.C. Lopez, M. Sordo, G. Elizondo and P. Ostrosky-Wegman, 2001. DNA breakage due to metronidazole treatment. Mutat. Res., 478: 153-158.
Direct Link - Miron, T., T. Bercovici, A. Rabinkov, M. Wilchek and D. Mirelman, 2004. Allicin: Preparation and applications. Anal. Biochem., 331: 364-369.
Direct Link - Morrison, R.N., P.B.B. Crosbie and B.F. Nowak, 2004. The induction of laboratory-based amoebic gill disease revisited. J. Fish Dis., 27: 445-449.
Direct Link - Munday, B.L., D. Zilberg and V. Findlay, 2001. Gill disease of marine fish caused by infection with Neoparamoeba pemaquidensis. J. Fish Dis., 24: 497-507.
Direct Link - Parsons, H., B. Nowak, D. Fisk and M. Powell, 2001. Effectiveness of commercial freshwater bathing as a treatment against amoebic gill disease in Atlantic salmon. Aquaculture, 195: 205-210.
Direct Link - Powell, M.D. and S.F. Perry, 1998. Acid-base and ionic fluxes in rainbow trout (Oncorhynchus mykiss) during exposure to chloramine-T. Aquat. Toxicol., 43: 13-24.
CrossRefDirect Link - Powell, M.D., D. Fisk and B.F. Nowak, 2000. Effects of graded hypoxia on Atlantic salmon infected with amoebic gill disease. J. Fish Biol., 57: 1047-1057.
Direct Link - Powell, M.D. and G.A. Clark, 2003. Efficacy and toxicity of oxidative disinfectants for the removal of gill Amoebae from the gills of amoebic gill disease affected Atlantic salmon (Salmo salar L.)in freshwater. Aquaculture, 220: 135-144.
Direct Link - Powell, M.D. and B.F. Nowak, 2003. Acid-base and respiratory effects of confinement in Atlantic salmon affected with amoebic gill disease. J. Fish Biol., 62: 51-63.
Direct Link - Powell, M., T. Green and C. Plaisted, 2003. A cell culture assay for the in vitro efficacy testing of amoebocidal compounds against Neoparamoeba pemaquidensis. Bull. Eur. Assoc. Fish. Pathol., 23: 291-296.
Direct Link - Re, J.L., M.P. De Meo, M. Laget, H. Guiraud, M. Castegnaro, P. Vanelle and G. Dumenil, 1997. Evaluation of the genotoxic activity of metronidazole and dimetridazole in human lymphocytes by the comet assay. Mutat. Res., 357: 147-155.
PubMedDirect Link - Roberts, S.D. and M.D. Powell, 2003. Reduced total hardness of freshwater enhances the efficacy of bathing as a treatment for amoebic gill disease in Atlantic salmon, Salmo salar L. J. Fish Dis., 26: 591-599.
Direct Link - Tona, L., K. Kambu, N. Ngimbi, K. Cimanga and A.J. Vlietinck, 1998. Antiamoebic and phytochemical screening of some congolese medicinal plants. J. Ethnopharmacol., 61: 57-65.
CrossRefDirect Link - Udoh, P. and A. Kehinde, 1999. Studies on antifertility effect on pawpaw seeds (Carica papaya) on the gonads of male albino rats. Phytother. Res., 13: 226-228.
Direct Link - Wen, G.Y., A. Mato, H.M. Wisniewski, M.N. Malik, E.C. Jenkins, A.M. Sheikh and K.S. Kim, 2004. Light and electron microscopic immunocytochemical localization of two major proteins in garlic bulb. J. Cell. Biochem., 58: 481-489.
Direct Link - Zilberg, D., V.L. Findlay, P. Girling and B.L. Munday, 2000. Effects of treatment with levamisole and glucans on mortality rates in Atlantic salmon (Salmo salar L.) suffering from amoebic gill disease. Bull. Eur. Assoc. Fish. Pathol., 20: 23-26.
Direct Link