
Preecha Prathepha
Rice Genetics Research Unit, Walai Rukhavej Botanical Research Institute (WRBRI),
Mahasarakham University, Khamriang District, Maha Sarakham Province, 44150, Thailand
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 26-33







ABSTRACT
The two wild rice species, Oryza rufipogon and O. nivara, which inhabit Thailand and the Lao PDR, are threatened with the loss of their natural habitats. The losses are primarily attributable to human intervention. Before this process advances, it is crucial to obtain basic information on the genetic variations of these species, so important knowledge about the current status of genetic variability. In this study, genetic variation within and between fourteen natural populations of the two populations from Thailand and Lao PDR was investigated at the DNA level by analysis of Polymerase Chain Reaction (PCR) mediate molecular markers. The results illustrated that wild rice from Thailand carried deletion (D) type ORF100, but deletion and non-deletion type (ND) were found in wild rice samples from the Lao PDR. Five different plastid subtypes (7C7A, 6C8A, 7C6A, 6C7A and 9C7A) were found in the collected samples. Both polymorphism and pattern of distribution of p-SINE1-r2 in the two wild rice species were found among the populations. These data indicated that genetic variation existed in these natural populations of wild rice, suggesting that the strategies should be developed that are conducive to the conservation of wild rice in its natural environment.
PDF Abstract XML References Citation
How to cite this article
Preecha Prathepha, 2008. Genetic Variation of Wild Rice Populations from Thailand and the Lao PDR Based on Molecular Markers. Pakistan Journal of Biological Sciences, 11: 26-33.
DOI: 10.3923/pjbs.2008.26.33
URL: https://scialert.net/abstract/?doi=pjbs.2008.26.33
DOI: 10.3923/pjbs.2008.26.33
URL: https://scialert.net/abstract/?doi=pjbs.2008.26.33
INTRODUCTION
Oryza sativa, an Asian cultivated rice, is now grown worldwide. It originated from a common ancestor with the AA genome. There are two species of the AA genome, O. nivara and O. rufipogon; each has been recognized as a common ancestor of the Asian cultivated rice. They are wild and occur throughout the tropics in places like the lowland river basin of the Amazon, Niger, Ganges-Bhramaputra, Mekong, the islands of Sri Lanka, Madagascar and New Guinea (Vaughan and Sitch, 1991). They play an important role as a valuable genetic resource and their continued availability remains crucial to high yield rice breeding programs (Xiao et al., 1998; Tian et al., 2006).
Thailand is considered to be one of the world`s richest rice genetic resource areas. Its biodiversity in both cultivated and wild rice is remarkably robust. Nonetheless, there have been relatively few attempts undertaken to characterize the wild rice gene pool, on the basis of genetic evaluation, primarily because information about the Indica-japonica differentiation is rarely available from natural genetic resources. Previous studies have examined genetic differentiation in only a limited number of wild rice strains (n <10) from Thailand (Sun et al., 2002). Quite simply, this means that the number of strains of wild rice used in these studies was insufficient to be able to discuss with any certainty the gene pool of these two ecotype strains in Thailand. Recently, wild rice with the AA genome from the Mekong River Basin was examined using molecular markers (Yamanaka et al., 2003). A few individuals in the examined sample were from Thailand. However, in the absence of genetically based identification, the genetic framework of these populations could not be fully understood. A genetically based identification of these rice is required if the populations genetic framework is to be understood. The ability to accurately compose an identification system is being compromised by the loss or alteration of arable systems, non-sustainable land use, deforestation and agro ecosystem deterioration, all of which have led to the loss of Thai wild rice populations (Chitrakon et al., 1992). Akimoto et al. (1999) suggested that the conservation of the wild rice population in Thailand should receive urgent attention in that the natural populations were dwindling in their natural habitats.
Thailand is uniquely rich in the wild relatives of Asian cultivated rice, which comprise an important part of the entire Thai rice gene pool. As reported in other studies, genetic impoverishment of high yielding rice cultivars is utilized during the breeding of genetic variations that are found in the natural populations of wild rice. For example, improving rice genetics for grain quality appears to be exploiting a novel gene from the origins of genetic diversity (Septiningsih et al., 2003). In Thailand, however, the influence of human activities on wild rice mainly includes land transformation, rice production, grazing and mowing, all of which have led to habitat destruction or fragmentation. These harmful activities have been ongoing since 1984, as reported by Morishima and Oka (1995) and continue to present day. Consequently, most wild rice localities have been transformed into cultivated rice fields, residential areas, factories and highways.
It is universally agreed that the primary solution to the genetic impoverishment of crop germplasm is genetic conservation and utilization during breeding of the vast genetic variations found among the natural populations of the wild progenitors and land races of cultivated plants (Tanksley and McCouch, 1997). The loss of wild rice populations has swiftly accelerated under human pressure and the changing agro ecosystems of Thailand. The loss is leading to a severe erosion of genetic diversity. Efforts to determine the genetic diversity of the wild rice gene pool serve not only to improve our understanding of the evolution, domestication and phylogeography of Asian cultivated rice, but also furnish a foundation for future worldwide rice breeding programs. The population structure of domesticated species is influenced by the natural history of the populations of its pre-domesticated ancestors, as well as by the breeding systems and complexity of the breeding practices exercised by humans (Garris et al., 2005). In addition, a better understanding of the genetics of wild rice and the association of some molecular markers with other cultivated/weedy traits will provide important information for the better management of both the negative and positive features associated with wild rice.
The purpose of this study is to establish a population genetic framework for the evaluation of wild rice species with the AA genome by characterizing the genetic differentiation of a collection of wild rice using plastid subtype ID (PS-ID) sequences based on the polymorphic DNA sequences of the linker sequences (Nakamura et al., 1998), plastid DNA marker (the ORF100 region of rice chloroplast DNA) and the p-SINE1-r2 that was found in the intron 10 of the rice waxy gene. The specific goals of this study are to report on the surveying, collection and characterization of the population structure of wild rice with the AA genome. The resulting framework will pose questions about the current status of genetic variability as it is inferred by the molecular marker technique used in genetic analysis. A large number of samples, collected from representative populations in all regions of Thailand, were used in this analysis.
MATERIALS AND METHODS
Plant materials: A total of 143 strains of the two wild rice species, O. nivara (n = 31) and O. rufipogon (n = 112) were collected from Thailand and Lao PDR and used in this study. In-field explorations of O. nivara populations were done in 2005-2006. This species is an annual and is found in shallow road-side ditches, where the land is completely desiccated in the dry season of Thailand and Lao PDR (March to May). O. rufipogon, on the other hand, is a perennial that inhabits either stable water conditions throughout the year, such as swamps, big and deep ponds, or places where there are unstable water conditions.
In the latter condition, O. rufipogon would propagate using its vegetative parts to produce a newly generated plantlet. The collection sites are shown in Table 1. Fresh leaves of single plants were collected from different plants which were well separated so as to represent genets. Samples of each population consisted of 2-23 plants. In total, 112 strains were examined for polymorphism of p-SINE1-r2 at the Waxy locus. Among them, 110 strains that represent all populations were next examined for polymorphism of the ORF100 region in the cpDNA. In addition, 25 strains of the two wild rice species were sampled from the total sample and examined for plastid subtype ID sequences of the linker sequences between rice plastid genes rpl16 and rpl14 (Nakamura et al., 1998).
DNA extraction, DNA markers analysis: Total genomic DNA was extracted by the CTAB method (Doyle and Doyle, 1987) from leaf tissues. To detect the ORF100 region of rice chloroplast DNA, a new primer set was designed to amplify the ORF100 region in the rice plasmid DNA, ORF100-f (5`-atgaaattgtataagtgg-3`)) and ORF100-r (5`-cagccgaggtcgtggtaaatcc-3`)). The INDEL marker can discriminate indica-japonica differentiation (Ishikawa et al., 2006). The polymerase chain reaction (PCR) was carried out using 0.5 units of Taq DNA polymerase, 0.5 μg total DNA, 10 pmol of each primer, 1.5 mM MgCl2, 0.1 mM dNTPs in a final volume of 20 μL. The PCR reactions were performed using the following profile: 94°C, 1 min; 50°C, 1 min and 72°C, 1.5 min for 35 cycles and a final extension of 5 min at 72°C. After PCR, the amplified products were electrophoresed for 45 min at 75 V and stained with ethidium bromide. The plasmid marker was classified into Deletion (D) and Non-Deletion (ND) types. The rice strain with the D type had 69 bp deletion in ORF100, while the rice with the ND type possessed 69 bp in the region (Kanno et al., 1993). The PCR products showing D type and/or ND type of some rice strains were further analyzed to confirm that they were the target amplified fragment of the direct sequencing of PCR products. The PCR products were analyzed on an ABI PRISM® 310 Genetic Analyzer (Applied Biosystems, Foster City, CA). To detect polymorphism of the linker sequences between the two rice plastid genes, PCR amplification of the plastid DNA fragment containing rpl16 and rpl14 was performed using a primer pairs (forward primer: 5`-aaagatctagatttcgtaaacaacatagaggaagaa-3`; reverse primer: 5`-atctgcagcatttaaaagggttctgaggttgaatcat-3`) (Nakamura et al., 1998). The reaction mixture containing 10 picograms of each primer, 0.2 mM of each dNTPs, 2 mM MgCl2, 0.5 units of Taq polymerase in a final volume of 20 μL. The amplification conditions were 45 cycles of 98°C for 10 sec, 55°C for 30 sec and 72°C for 1 min. A final extension step at 72°C for 5 min was performed after the 45 cycles. PCR products were resolved using 2.0% agarose gel electrophoresis. The purified PCR products were used as a template DNA for direct sequencing as described above.
Polymerase chain reaction and nested PCR, to detect the presence or absence of p-SINE1-r2 at the waxy locus, were carried out using protocols described by previous researchers (Yamanaka et al., 2003). A primer set, Wax-2F (5` acgccggtggagggcaggaagatcaact-3`) and Wax-4R (5`-tcaaggagcagccacgttctccttggcg-3`) were used to amplify the second half of the rice waxy gene. For the nested PCR, a primer set, M6 ((5`-ggaggacgtgc agatcgttc-3`) and R7 ((5`- acgagtccaccggtg gacgc-3`) designed by Umeda et al. (1991), was used to amplify the expected fragment containing p-SINE1-r2 as reported previously (Hirano et al., 1994). PCR amplification and gel electrophoresis using the two primer sets were performed under the same condition as described by Yamanaka et al. (2003).
RESULTS
Polymorphism of the intron 10 of Wx gene in wild rice populations: As shown in Fig. 1, polymorphism at the intron 10 of Wx gene, revealed by the presence or absence of the p-SINE1-r2, was detected by PCR. Of the 112 strains of O. rufipogon examined, three showed patterns of polymorphism. Sixty-two out of 112 (55.4%) strains were heterozygous (+/-) in this region, 44 strains (39.2%) did not possess p-SINE1-r2 and six strains (5.4%) possessed this element (+/+). Meanwhile, 31 strains of O. nivara examined showed three patterns of polymorphism. Nineteen out of 31 (61.3%) strains of O. nivara contained the p-SINE1-r2 (+/+) and 11 strains (35.5%) did not (-/-) and only one strain (3.2%) was heterozygous (Table 1). For O. rufipogon, several
| Table 1: | Study sites of Oryza rufipogon (OR) and O. nivara (ON) and the distribution of p-SINE1-r2 in each population |
 | |
 | |
| Fig 1: | PCR analysis of the p-SINE1-r2 in intron 10 of the rice waxy gene of wild rice strains examined (lanes No. 1-47). Polymorphism is caused by insertion (+) or non-insertion (-) of the element are observed. Genotypes are indicated by (+/+), (+/-) and (-/-) for the upper band (insertion), heterozygote and lower band (non-insertion), respectively. M = molecular ladders (1 kb DNA ladder, Gibco, BRL). For example, lanes No. 2-25 and lanes No. 29-34 show homozygote (+/+) and (-/-), respectively. Lanes No. 28, 36-43 and 46 are heterozygote (+/-) |
 |
 | |
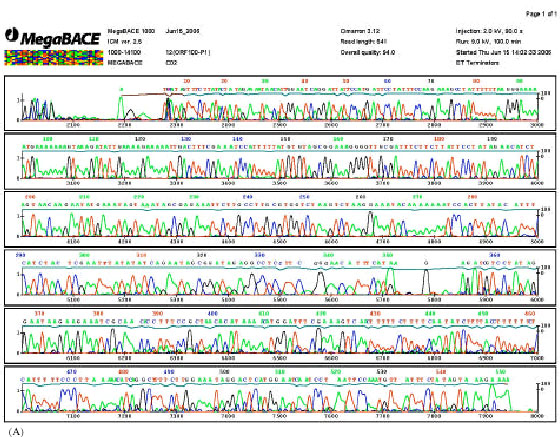
| Fig 2: | The electropherograms showed deletion type (D) (A) and non-deletion type (ND) (B) within the ORF100 region of chloroplast DNA in wild rice examined. Direct repeat of DNA sequence, GAAAAAAAGTAA, as shown in the upstream end (90-108 bp) and down stream end (175-186 bp) used as DNA marker for ND type compares to D type |
populations showed polymorphism within the population, whereas some populations were monomorphic, i.e., Ban Klang Noi, Nakhon Phanom Province, Muang District Chiang Rai Province, Kosum Pisai District, Maha Sarakham Province and Ban Keon, Vientiane, Lao PDR.
Chloroplast DNA (cpDNA) polymorphism in wild rice: Cytoplasmic markers were determined by the polymorphism of the ORF100 region in the cpDNA and the linker sequences between rice plastid genes rpl16 and rpl14 (Fig. 2,3). A wild relative, O. rufipogon from Thailand, has been documented for ORF100 polymorphism (Chen et al., 1992). Both types, (D), the deletion of the 69 base pairs of the ORF 100 region of the cpDNA and (ND), without the deletion of the 69 base pairs of the ORF 100 region, were found in the samples. All wild rice strains (n = 83) of O. rufipogon from Thailand were D type, whereas 16 out of 27 (59.3%) of this wild rice species from Lao PDR showed D type; the 11(40.7%) remaining were ND type. This suggests that the wild rice from Lao PDR showed ORF100 polymorphism in the population used in this study.
Polymorphism of plastid DNA has been reported for Thai wild rice strains (Nakamura et al., 1998). In the present study, plastid subtype sequences were examined for 31 strains of both types of randomly chosen wild rice. These wild rice samples showed five different plastid subtypes (7C7A, 6C8A, 7C6A, 6C7A and 9C7A). O. nivara (n = 9) showed two plastid subtypes (7C6A and 7C7A); whereas, O. rufipogon showed five plastid subtypes as mentioned (Table 2). Polymorphism of the plastid subtype was found within and/or among populations. The Chiang Rai population showed 7C7A and 6C8A; whereas, two populations of Vientiane, Lao PDR revealed 7C6A, 6C7A and 6C8A, respectively. The 9C7A plastid subtype was found only in strains examined of O. rufipogon from the Sakon Nakhon population
| Table 2: | Chloroplast DNA markers for Oryza rufipogon (OR) and O. nivara (ON) from Thailand and Lao PDR |
 | |
*D: The deletion of the 69 base pairs of the ORF 100 region of the cpDNA and ND, without the deletion of the 69 base pairs of the region, **: A linker sequence between rpl16 and rpl14 plastid genes | |
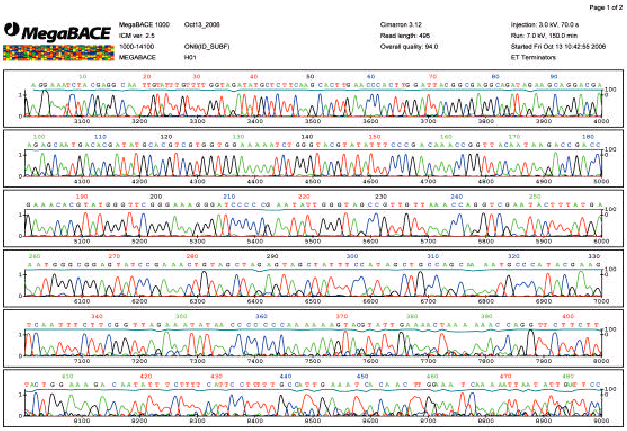 | |
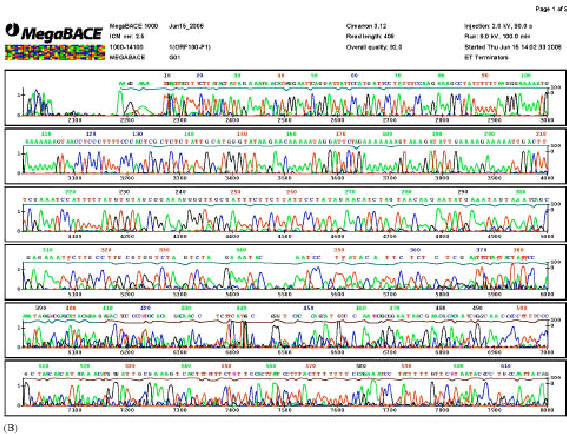
| Fig 3: | Polymorphism of the linker sequence between rice plastid genes rpl16 and rpl14 is observed in wild rice strain analyzed. This electropherogram reveals genotype 7C6A at the position 350-370 bp of DNA sequence |
DISCUSSION
The use of molecular markers is considered best for analysis of genetic diversity and variety identification because there is no effect on the stage of development, environment or management practices. Among the various classes of molecular markers, the genetic differentiation in rice strains was investigated by detecting the 69 bp deletion of the ORF100 region of chloroplast DNA (Kanno et al., 1993; Chen et al., 1993) and by analysis of the plastid subtype (PS-ID) sequence (Ishikawa et al., 2006). In addition, genetic variability at the Waxy locus has been used extensively for the identification and characterization of rice accessions (Yamanaka et al., 2003; Prathepha and Baimai, 2004), relationship with amylose content (Bergman et al., 2001) and identification of glutinous rice accessions (Wanchana et al., 2003).
There is very little information available on the nature and extent of the genetic diversity of the wild rice of Thailand, particularly information based on molecular markers. This kind of information would be valuable for the rationalization of wild rice germplasm conservation and the utilization of the germplasm in a breeding program. Hence, the present study was conducted in an attempt to understand the pattern of genetic variability in large wild rice accessions based on molecular markers.
Chloroplast DNA polymorphism in wild rice population: Genetic variability in wild rice accessions has been previously documented (Chen et al., 1992; Akimoto et al., 1999; Yamanaka et al., 2003; Nakamura et al., 1998). The present study combined a large number of wild rice accessions (n = 143) and these accessions showed significant differentiation within and/or among populations. The genetic structure obtained from this study is a legacy of structure in ancestral rice populations. In an analysis of polymorphism in the ORF100 region from Thailand, no variation in the ORF100 region was found. The sampled accessions were monomorphic (deletion type), suggesting that there is genetic similarity (indica type) found in Thai wild rice with the AA genomes used in this study. The results from this study support the hypothesis that the beginnings of indica cultivars were in the flooded plains of large rivers, such as the Mekong in Laos-Cambodia-Vietnam and the Chaopraya in Thailand (Sato, 1996). In addition, a few strains examined from Vientiane, Lao PDR showed both D (indica type) and ND (japonica type) types, supporting the hypothesis that japonica and indica cultivated rice are derived from independent domestication events from ancestral rice that had already differentiated into two gene pools and had different maternal origins (Cheng et al., 2003; Londo et al., 2006). Indeed, japonica rice cultivars are found mostly in the northern region of Thailand, whereas most of the upland rice cultivars grown in the highlands and mountainous regions of northern Laos have been classified as japonica rice (Yamanaka et al., 2001).
Nakamura et al. (1998) reported that four chloroplast subtypes, 6C7A, 6C8A, 7C7A and 9C7A, were found in O. nivara, while three subtypes, 6C7A, 7C6A and 6C8A were found in O. rufipogon. All of these subtypes were found in the wild rice samples of this present study. The plastid subtype 8C8A was not found in either study. With respect to the chloroplast subtypes, they are of interest for cultivated rice evolutionary reasons. The variation of subtypes found in rice cultivars reflects their different origins. The data suggest that the Indica-japonica differentiation occurs not only in cultivated rice but also in its wild relatives. The data further suggest that indica and japonica types belong to different chloroplast types. These results indicate that the Indica-japonica differentiation of wild rice may have occurred before domestication of cultivated rice. This information is valuable for an understanding of rice evolution and supports previous studies as mentioned above.
Polymorphism of nuclear DNA marker, p-SINE1-r2, in wild relatives of cultivated rice: The wild relatives of cultivated rice (O. sativa L.) have been documented for polymorphism of p-SINE1-r2 (Yamanaka et al., 2003). The Yamanaka study was designed to address the association between the p-SINE1-r2 polymorphism and the annual type (O. nivara) and the perennial type (O. rufipogon). The study showed that the p-SINE1-r2 polymorphism was highly associated with the annual-perennial differentiation. In addition, the study attempted to propose a new collection method for materials of wild rice, particularly for the perennial type. Interestingly, three type of polymorphism (-/-, +/- and +/+) were found in the wild relatives of the perennial type which were collected from Thailand. Of the samples examined in this present study, only 6 (4.2%) wild rice strains of the perennial type showed the genotype (+/+). These 6 individuals were collected from a population growing next to a rice paddy field at Vientiane, Laos PDR. This genotype was not found in the Thai wild relatives of the perennial type. All the annual types (O. nivara) possessed p-SINE1-r2 resulting in these individuals having the genotype (+/+). Regardless of the problems associated with collection methods, the perennial type that carried the genotype (+/+) may have been caused by a gene flow from cultivated rice to wild rice as reported by Chen et al. (2004) and Song et al. (2003). The reasoning which supports this idea is the genotype (+/+) was the prominent genotype of cultivated rice. All rice cultivars examined for the p-SINE1-r2 polymorphism showed only the genotype (+/+) (Prathepha, 2003). However, another possible explanation is that the original population of wild relatives in Thailand exhibited prominent genotypes (-/-) and (-/+) and mainly propagated by vegetative reproduction. Consequently, these genotypes were detected in all populations of Thai wild relatives. Individuals carrying the genotype (+/+) were not found in the wild rice strains examined in this study. In an evolutionary context, many of the most intriguing questions remain to be answered, such as what are the effects that influenced genotypic distribution and/or genotypic survival in the natural habitat of the genotype (+/+) in wild rice. Studies designed to address these and other questions will lead to a better understanding of the adaptation of O. rufipogon and processes of domestication. This study revealed the existence of considerable genetic variation in the wild rice of Thailand and Lao PDR. Knowledge about the distribution of genetic variation among or within populations of wild rice plays a crucial role in their conservation. For natural locations, these types of data can assist in determining where and how to sample wild rice populations in order to optimize diversity captured for ex situ population. The genetic erosion of wild rice in Thailand is caused not only by the disturbance of their natural habitats by various human activities but also by gene flow from cultivated rice to wild rice (Akimoto et al., 1999). Gene flow from cultivated rice to wild rice may effect the dynamics of the wild rice population. The structure of the wild rice population, as it is affected by cultivated rice, is a core issue for the biosafety of the wild rice gene pool, particularly since there are many worldwide programs involved in the development of transgenic rice. When alien transgenes escape to wild rice, they may persist and disseminate within the weedy or wild populations through sexual reproduction and/or vegetative propagation. The escape of these transgenes will likely cause ecological problems. This interspecific hybridization, a factor affecting the genetic structure of wild rice, has been reported by Chen et al. (2004) and Kuroda et al. (2005). Consequently, the aggressive spread of hybrids with superior ecological stamina could lead to the extinction of populations of wild rice species in local ecosystems (Ellstrand and Elam, 1993). Although GM rice is not grown in Thailand, it nonetheless should remain a concern of policy makers. In addition, some traits, which are unique to wild rice, might be beneficial to the improvement of cultivated rice. In Thailand, strategies should be developed that are conducive to the conservation of wild rice in its natural environment.
ACKNOWLEDGMENTS
The author sincerely appreciates P. Dulfer for his continuous encouragement. This research was supported in part by the grant from Mahasarakham University.
REFERENCES
- Akimoto, M., Y. Shimamoto and H. Morishima, 1999. The extinction of genetic resources of Asian wild rice, Oryza rufipogon Genet. Resour. Crop Evol., 46: 419-425.
CrossRef - Bergman, C.J., J.T. Delgado, A.M. McClung and R.G. Fjellstrom, 2001. An improve method for using a microsatellite in the rice waxy gene to determine amylose class. Cereal Chem., 78: 257-260.
CrossRefDirect Link - Chen, W.B., I. Kakamura, Y.I. Sato and H. Nakai, 1992. Distribution of different chloroplast DNA types in Indica and japonica rice. Rice Genet. Newslett., 9: 142-144.
Direct Link - Chen, W.B., I. Nakamura, Y.I. Sato and H. Nakai, 1993. Distribution of deletion type in cpDNA of cultivated and wild rice. Jap. J. Genet., 68: 597-603.
Direct Link - Chen, L.J., D.S. Lee, Z.P. Song, H.S. Suh and B.R. Lu, 2004. Gene flow from cultivated rice (Oryza sativa) to its weedy and wild relatives. Ann. Bot., 93: 67-73.
Direct Link - Cheng, C., R. Motohashi, S. Tsuchimoto, Y. Fukuta, H. Ohtsubo and E. Ohtsubo, 2003. Polyphyletic origin of cultivated rice: Based on the interspersion pattern of SINEs. Mol. Biol. Evol., 20: 67-75.
Direct Link - Chitrakon, S., Y.I. Sato, H. Morishima and Y. Shimamoto, 1992. Genetic erosion of rice in Thailand. Rice Genet. Newslett., 9: 73-74.
Direct Link - Doyle, J.J. and J.L. Doyle, 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull., 19: 11-15.
Direct Link - Ellstrand, N.C. and D.R. Elam, 1993. Population genetic consequences of small population size-implications for plant conservation. Ann. Rev. Ecol. Syst., 24: 217-242.
CrossRefDirect Link - Garris, A.J., T.H. Tai, J. Coburn, S. Kresovich and S. McCouch, 2005. Genetic structure and diversity in Oryza sativa L. Genetics, 169: 1631-1638.
CrossRefPubMedDirect Link - Hirano, H.Y., K. Mochizuki, M. Umeda, H. Ohtsubo, E. Ohtsubo and Y. Sano, 1994. Retrotransposition of a plant SINE into the wx locus during evolution of rice. J. Mol. Evol., 38: 132-137.
PubMedDirect Link - Ishikawa, R., S. Yamanaka, Y. Fukuta, S. Chitrakon and C. Bounphanousay et al., 2006. Genetic erosion from modern varieties into traditional upland rice cultivars (Oryza sativa L.) in Northern Thailand. Genet. Resour. Crop Evol., 53: 245-252.
CrossRefDirect Link - Kanno, A., N. Watanabe, I. Nakamura and A. Hirai, 1993. Variations in chloroplast DNA from rice (Oryza sativa): Differences between deletions mediated by short direct-repeat sequences within a single species. Theor. Applied Genet., 86: 579-584.
CrossRefDirect Link - Kuroda, Y., Y.I. Sato, C. Bounphanousay, Y. Kono and K. Tanaka, 2005. Gene flow from cultivated rice (Oryza sativa L.) to wild Oryza species (O. rufipogon Griff. and O. nivara Sharma and Shastry) on the Vientiane plain of Laos. Euphytica, 142: 75-83.
Direct Link - Londo, J.P., Y.C. Chiang, K.H. Hung, T.Y. Chiang and B.A. Schaal, 2006. Phylogeography of Asian wild rice, Oryza rufipogon, reveals multiple independent domestications of cultivated rice, Oryza sativa. Proc. Nat. Acad. Sci. USA., 103: 9578-9583.
Direct Link - Morishima, H. and H.I. Oka, 1995. Genetic erosion in wild and cultivated rice species. Rice Genet. Newslett., 12: 168-170.
Direct Link - Nakamura, I., H. Urairong, N. Kameya, Y. Fukuta, S. Chitrakon and Y.I. Sato, 1998. Six different plastid subtype were found in O. sativa, O. rufipogon complex. Rice Genet. Newslett., 15: 80-83.
Direct Link - Prathepha, P., 2003. Distribution of p-SINE1-r2 at the Wx locus in cultivated and wild rice in Thailand. IRRN., 28.1: 26-27.
Direct Link - Prathepha, P. and V. Baimai, 2004. Variation of wx microsatellite allele, waxy allele distribution and differentiation of chloroplast DNA in a collection of Thai rice (Oryza sativa L.). Euphytica, 140: 231-237.
Direct Link - Septiningsih, E.M., J. Prasetiyono, E. Lubis, T.H. Tai, T. Tjubaryat, S. Moeljopawiro and S.R. McCouch, 2003. Identification of quantitative trait loci for yield and yield components in an advanced backcross population derived from the Oryza sativa variety IR64 and the wild relative O. rufipogon. Theor. Applied Genet., 107: 1419-1432.
Direct Link - Song, Z.P., B.R. Lu, Y.G. Zhu and J.K. Chen, 2003. Gene flow from cultivated rice to the wild species Oryza rufipogon under experimental field conditions. New Phytol., 157: 657-665.
Direct Link - Sun, Q., K. Wang, A. Yoshimura and K. Doi, 2002. Genetic differentiation for nuclear, mitochondrial and chloroplast genomes in common wild rice (Oryza rufipogon Griff.) and cultivated rice (Oryza sativa L.). Theor. Applied Genet., 104: 1335-1345.
Direct Link - Tanksley, S.D. and S.R. McCouch, 1997. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science, 277: 1063-1066.
CrossRefPubMedDirect Link - Tian, F., D.J. Li, Q. Fu, Z.F. Zhu, Y.C. Fu, X.K. Wang and C.Q. Sun, 2006. Construction of introgression lines carrying wild rice (Oryza rufipogon Griff.) segments in cultivated rice (O. sativa L.) background and characterization of introgressed segments associated with yield-related traits. Theor. Applied Genet., 112: 570-580.
Direct Link - Umeda, M., H. Ohtsubo and E. Ohtsubo, 1991. Diversification of the rice Waxy gene by insertion of mobile DNA elements into introns. Jap. J. Genet., 66: 569-586.
CrossRef - Vaughan, D.A. and L.A. Sitch, 1991. Gene flow from the jungle to farmers. Wild rice genetic resources and their uses. BioScience, 41: 22-28.
Direct Link - Wanchana, S., T. Toojinda, S. Tragoonrung and A. Vanavichit, 2003. Duplicated coding sequence in the waxy allele of tropical glutinous rice (Oryza sativa L.). Plant Sci., 165: 1193-1199.
Direct Link - Xiao, J., J. Li, S. Grandillo, S.N. Ahn, L. Yuan, S.D. Tanksley and S.R. McCouch, 1998. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics, 150: 899-909.
Direct Link - Yamanaka, S., I. Nakamura, H. Nakai and Y.I. Sato, 2003. Dual origin of the cultivated rice based on molecular markers of newly collected annual and perennial strains of wild rice species, Oryza nivara and O. rufipogon. Genet. Resour. Crop Evol., 50: 529-538.
Direct Link