
A. Ghasemi
Department of Pathobiology, School of Public Health, Medical Sciences/University of Tehran, Tehran, Iran
M.H. Shirazi
Department of Pathobiology, School of Public Health, Medical Sciences/University of Tehran, Tehran, Iran
R. Ranjbar
Molecular Biology Research Center, Baqiyatallah University of Medical Sciences, Tehran, Iran
M.R. Khorramizadeh
Department of Biotechnology, School of Public Health, Medical Sciences/University of Tehran, Tehran, Iran
N.E. Daryani
Faculty of Medicine, Medical Sciences/University of Tehran, Tehran, Iran
M. Hosseini
Department of Biostatistics, School of Public Health, Medical Sciences/University of Tehran, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 22 | Page No.: 2579-2583







ABSTRACT
The aim of this study was to investigate the prevalence of cagA and cagE genes in H. pylori strains isolated from different patient groups with Non-Ulcer Dyspepsia (NUD), Duodenal Ulcer (DU), Gastric Ulcer (GU) and Gastric Cancer (GC). The patients admitted to the gastroenterology unit at Sharyati hospital in Tehran in 2006 were included in this study. Gastric biopsy specimens were obtained from the antrum of the stomach from each patient then cultured for detection of H. pylori. Identification of H. pylori was performed according to the standard bacteriological methods. Genomic DNA was extracted using a commercially available Qia gene kit. PCR was done using primers cagA-F, cagA-R and cagE-F, cagE-R to detect the target genes cagA and cagE, respectively. Amplified products of target genes were confirmed by sequencing. The cagA and cagE were detected among 85 and 86% of H. pylori isolates, respectively. Prevalence of cagA and cagE genes in the patients with NUD, DU, GU and GC were 22 (64.7%), 28 (100%), 18 (90%), 10 (100%) and 25 (73.5%), 27 (96.4%), 19 (95%), 7 (70%), respectively. The current study demonstrated a significant correlation between peptic ulceration and the presence of H. pylori isolates carrying cagE and cagA genes in Iranian patients.
PDF Abstract XML References Citation
How to cite this article
A. Ghasemi, M.H. Shirazi, R. Ranjbar, M.R. Khorramizadeh, N.E. Daryani and M. Hosseini, 2008. The Prevalence of CagA and CagE Genes in Helicobacter pylori Strains Isolated from Different Patient Groups by Polymerase Chain Reaction. Pakistan Journal of Biological Sciences, 11: 2579-2583.
DOI: 10.3923/pjbs.2008.2579.2583
URL: https://scialert.net/abstract/?doi=pjbs.2008.2579.2583
DOI: 10.3923/pjbs.2008.2579.2583
URL: https://scialert.net/abstract/?doi=pjbs.2008.2579.2583
INTRODUCTION
Gastroenteritis is a common and important public health problem (Ranjbar et al., 2007a). Colonization of the stomach mucosa by H. pylori is a major cause of acute and chronic gastric pathologies in humans. Several virulence genes of H. pylori have been identified. The most important determinants are cagE and cagA genes in the cag pathogenicity island (cagPAI) genes and vacA (Tan et al., 2006). The cytotoxin associated gene A (cagA) provides a key marker for the cag-PAI present in type I strains and its product, the cagA protein, has been shown to be delivered into cultured gastric epithelial cells (AGS cells) and immediately phosphorylated close to the site of attachment of the bacteria (Argent et al., 2008; Odenbreit et al., 2001; Stein et al., 2000; Asahi et al., 2000; Backert et al., 2000). The type I isolates may differ in structure of the cag-PAI as proposed by Censini et al. (1996), but evidence from investigation of multiple loci suggest that most isolates contain an uninterrupted and intact cag-PAI (Jenks et al., 1998; Slater et al., 1999; Occhialini et al., 2001; Owen et al., 2001; Maeda et al., 1999). Cytotoxin associated gene E (cagE) is also one of the marker genes in cagI of the cag PAI. It is essential for cagA translocation and phosphorylation (Odenbreit et al., 2000). The presence of the cagE gene has been associated with a bad clinical outcome, especially in developed countries (Yamazaki et al., 2005). The vacuolating cytotoxin induces cytoplasmic vacuolation in a variety of mammalian cell lines in vitro and produces epithelial cell damage and mucosal ulceration when administered intragastrically in mice (Lin et al., 2000). However, there seems to be no functional link between cagA and vacA and it is likely that cagA is a genotypic marker for the presence and/or expression of other ulcer- or cancer-related virulence genes (Evans et al., 1998).
Molecular approach has provided powerful tools for diagnosis, epidemiological surveillance and tracking of key genes among the microbial pathogens (Ranjbar et al., 2007b). The aims of the present study were to determine the prevalence of cagA and cagE genes in Iranian patients with non-ulcer dyspepsia, duodenal ulcer, gastric ulcer and gastric cancer by PCR.
MATERIALS AND METHODS
In the present study, a total of 150 Iranian patients (78 male and 72 female; mean age 40.9 years; ranged from 16 to 79 years) admitted to the gastroenterology unit in Sharyati Hospital, Tehran, in the years 2005 and 2006 were enrolled for upper endoscopy. An informed consent was obtained from all patients who were included in the study according to the protocol approved by the local ethics committee.
| Table 1: | The primers set used in the study |
 | |
During endoscopic examination, gastric biopsy specimens were obtained from the antrum of the stomach. Gastric biopsy specimens from each patient were inoculated onto Brucella agar base medium containing sheep blood (10%) (Merck, Germany) and antibiotic supplement (Merck, Germany) and cultured for 3 to 5 days at 37°C under microaerobic conditions (5% O2, 5% CO2, 90% N2). Helicobacter pylori strains was identified by typical gram stain, colony morphology, and by positive biochemical tests for urease, catalase and oxidase (Smith et al., 2002).
The patients who infected with H. pylori were clustered into four groups according to their clinical and endoscopic presentation: non-ulcer dyspepsia (NUD; n = 34), duodenal ulcer (DU; n = 28), gastric ulcer (GU; n = 20) and gastric cancer (GC; n = 10).
Bacterial genomic DNA was extracted using a commercially available kit (Qia gene, Hilden, Germany) according to manufacturer`s instructions. PCR was used to detect the cagA, cagE genes. All primer sets used were selected from the published literature as shown in Table 1.
The master mixes used for PCR consisted of 5 mM of 10x PCR buffer, 500 mmol of KCl, 100 mmol of Tris-HCl (pH 8.8), 2 mM MgCl2, 250 μM each of the four deoxynucleoside triphosphates, 0.5 mM of each primer and 0.3 mM of Taq DNA polymerase (BioEngland). Five microliters from each H. pylori diluted extract, positive control DNA, or sterile water (extraction blank and negative control) was added to the mixture to obtain a final volume of 50 μL. PCR amplification was performed according to earlier reports (Stone et al., 1997; Tomasini et al., 2003). For cagA and cagE, PCR conditions were as follows: 3 min at 95°C and then 50 cycles of 94°C for 1 min, 48°C (cagA) and 53°C (cagE) for 45 sec and 72°C for 45 sec. PCRs were performed using a Robocycler Gradient 40 temperature cycler (Stratagene).
RESULTS AND DISCUSSION
A total of 92 H. pylori strains were isolated from different groups of patients: NUD (n = 34), DU (n = 28), GU (n = 20) and GC (n = 10).
| Table 2: | Prevalence of cagA and cagE genes in different patient groups |
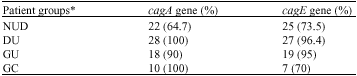 | |
| *NUD: Non-Ulcer Dyspepsia, DU: Duodenal Ulcer, GU: Gastric Ulcer, GC: Gastric Cancer | |
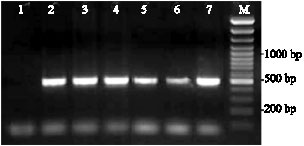 | |
| Fig. 1: | Detection of PCR products of cagA gene by agarose gel electrophoresis. Lane 1: Negative control, Lanes 2-6: Clinical representative samples, Lane 7: Positive control (ATCC 43504 strain), M: Molecular weight marker (100 bp) |
Overall, cagA was detected in 78 (85%) of the isolates. The carriage of cagA-positive strains of H. pylori in the patients with DU was 100% (28/28), with GU, 90% (18/20), with GC, 100% (10/10) and with NUD, 64.7% (22/34). A significant difference was observed in carriage of the cagA-positive strains of H. pylori in those with PUD and Gastric cancer compared to NUD (p<0.05) (Table 2).
The cagE positive H. pylori strains were isolated from 79 patients (86%). The frequency of cagE-positive strains in patients with PUD was 92% (46/48). Seventy percent and 73.5% of strains isolated from patients with GC and NUD were cagE and cage positive, respectively (Table 2). Figure 1 and 2 show the expected amplified fragments of cagA and cagE genes, respectively in PCR reaction. Amplified products of target genes were confirmed by sequencing. The confirmed sequences have been submitted to GenBank (accession numbers DQ512724 and DQ991147).
Helicobacter pylori infection is extremely common worldwide with a prevalence ranging from 25% in developed countries to more than 80% in the developing world (Parsonnet, 2003; Pounder and Ng, 1995). Various factors such as the environment, host genetic factors and bacterial virulent ability contribute to infection outcome caused by H. pylori (Campbell et al., 1997; Malaty and Graham, 1994 ).
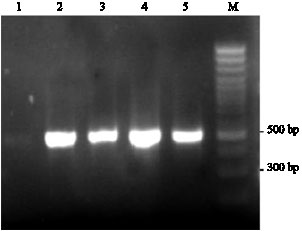 | |
| Fig. 2: | Identification of the cagE by Polymerase Chain Reaction (PCR). Lanes 1: Negative control sample (without DNA); Lanes 2-4, Clinical representative samples, Lane 5: Positive control H. pylori ATCC43504, M: Molecular weight marker (100 bp) |
The different molecular methods could provide sensitive interpreting keys suitable for microbiological studies (Ranjbar et al., 2008a, b). In present study we applied PCR to investigate the prevalence of cagA and cagE genes in Iranian patients with non-ulcer dyspepsia, duodenal ulcer, gastric ulcer and gastric cancer. We also studied the relationships between the presence of H. pylori strains carrying cagE and cagA genes and the clinical outcome in the patients studied.
A higher prevalence of the cagA gene was observed in the patients with DU (100%) and gastric cancer when compared to the NUD group (64.3%) (p<0.05).
Subsequent studies have shown more inconsistent results (Kim et al., 2004; Proença Módena et al., 2007). The current study demonstrated that the majority (85%) of H. pylori strains isolated from Iranian patients were cagA positive. This finding is similar to the pattern usually described in Asian populations (Hirata et al., 2004; Zhou et al., 2004). Oliveira et al. (2003) demonstrated that more than 79% of subjects with ulcer disease in Brazil were infected with H. pylori strains carrying cagA. They also showed that the prevalence of cagA in the patients with gastritis, duodenal ulcer and gastric carcinoma were 59.21, 90 and 94.23%, respectively. These data are similar to present findings in this study. In contrast, Aydin et al. (2004) reported that only 59.2% of Turkish strains carried cagA gene and prevalence of cagA in patients with PUD and NUD were 72.3 and 47%, respectively. In China and Japan, cagA-positive strains are nearly universally present and are not associated with disease complications (Hirata et al., 2004; Zhou et al., 2004).
It has been reported that infection with a cagE positive H. pylori strain is associated with the presence of duodenal ulcer. In addition, Day et al. (2000) reported that infection of gastric cells in tissue culture by cagE positive H. pylori resulted in greater increments in IL-8 levels compared with cagE-negative strains and concluded that enhanced chemokine production after infection with cagE-positive H. pylori could affect disease outcome for duodenal ulcer.
In this study, 96.4% (27/28) of strains isolated from patients with duodenal ulcer carried the cagE gene.
This study also demonstrated that infection with a cagE-positive H. pylori strain was associated with peptic ulcer disease 95.7% (46/48). Similarly, preliminary data from this study also show that the presence of the cagE gene in strains of H. pylori is associated with duodenal ulceration. For instance, in a study by Fallone et al. (2000) 31 (37%) of 84 patients with gastroduodenal disease (including both peptic ulceration and gastric cancer) were infected with cagE-positive strains, compared to only 20.7% of 92 patients with gastritis alone. However, association of cagE in patients with GC and NUD is equal in present study and it was consistent with other studies where no difference was found in the frequency of cagE positive isolates among patients with gastritis, duodenal ulcer or gastric cancer (Hsu et al., 2002; Tan et al., 2006).
In this study, 69 H. pylori strains carried both cagA and cagE genes and only 4 strains did not carry each of them. Seventy-four percent stains associated with PUD, carried both of cagA and cagE genes.
The current study demonstrated a significant correlation between peptic ulceration and the presence of H. pylori isolates carrying cagE and cagA genes in Iranian patients studied.
ACKNOWLEDGMENT
This research was supported in part by funds from Medical Sciences/University of Tehran, Tehran, Iran.
REFERENCES
- Argent, R.H., J.L. Hale, E.M. El-Omar and J.C. Atherton, 2008. Differences in Helicobacter pylori CagA tyrosine phosphorylation motif patterns between western and East Asian strains and influences on interleukin-8 secretion. J. Med. Microbiol., 57: 1062-1067.
CrossRefPubMedDirect Link - Asahi, M., T. Azuma, S. Ito, Y. Ito and H. Suto et al., 2000. Helicobacter pylori cagA protein can be tyrosine phosphorylated in gastric epithelial cells. J. Exp. Med., 191: 593-602.
PubMedDirect Link - Aydin, F., N. Kaklikkaya, O. Ozgur, K. Cubukcu, A.O. Kilic, I. Tosun and M. Erturk, 2004. Distribution of vacA alleles and cagA status of Helicobacter pylori in peptic ulcer disease and non-ulcer dyspepsia. Clin. Microbiol. Infect., 10: 1102-1104.
CrossRefPubMedDirect Link - Backert, S., E. Ziska, V. Brinkmann, U. Arndt and A. Fauconnier et al., 2000. Translocation of the Helicobacter pylori cagA protein in gastric epithelial cells by a type IV secretion apparatus. Cell. Microbiol., 2: 155-164.
CrossRefPubMedDirect Link - Campbell, S., A. Fraser, B. Holliss, J. Schmid and P.W. O'Toole, 1997. Evidence for ethnic tropism of Helicobacter pylori. Infect. Immun., 65: 3708-3712.
PubMedDirect Link - Censini, S., C. Lange, Z. Xiang, J.E. Crabtree and P. Ghiara et al., 1996. Cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA., 93: 14648-14653.
PubMedDirect Link - Day, A.S., N.L. Jones, J.T. Lynett, H.A. Jennings, C.A. Fallone, R. Beech and P.M. Sherman, 2000. ca ge is a virulence factor associated with Helicobacter pylori induced duodenal ulceration in children. J. Infect. Dis., 181: 1370-1375.
PubMedDirect Link - Fallone, C.A., A.N. Barkun, M.U. Guttke, L.M. Best and V.G. Lo et al., 2000. Association of Helicobacter pylori genotype with gastroesophageal reflux disease and other upper gastrointestinal diseases. Am. J. Gastroenterol., 95: 659-669.
CrossRefPubMedDirect Link - Hirata, Y, A. Yanai, W. Shibata, Y. Mitsuno, S. Maeda, K. Ogura, H. Yoshida, T. Kawabe and M. Omata, 2004. Functional variability of cagA gene in Japanese isolates of Helicobacter pylori. Genetic, 343: 165-172.
CrossRefPubMedDirect Link - Hsu, P.I., I.R. Hwang, D. Cittelly, K.H. Lai and H.M.T. El-Zimaity et al., 2002. Clinical presentation in relation to diversity within the Helicobacter pylori cag pathogenicity Island. Am. J. Gastroenterol., 97: 2231-2238.
CrossRefPubMedDirect Link - Jenks, P.J., F. Megraud and A. Labigne, 1998. Clinical outcome after infection with Helicobacter pylori does not appear to be reliably predicted by the presence of any of the genes of the cag pathogenicity Island. Gut, 43: 752-758.
PubMedDirect Link - Kim, J.W., J. Gyukim, S.L. Chae, Y.J. Cha and S.M. Park, 2004. High prevalence of multiple strain colonization of Helicobacter pylori in Korean patients: DNA diversity among clinical isolates from the gastric corpus, antrum and duodenum. Korean J. Internal Med., 19: 1-9.
PubMedDirect Link - Maeda, S., H. Yoshida, T. Ikenoue, K. Ogura and F. Kanai et al., 1999. Structure of cag pathogenicity Island in Japanese Helicobacter pylori isolates. Gut, 44: 336-341.
PubMedDirect Link - Malaty, H.M. and D.Y. Graham, 1994. Importance of childhood socioeconomic status on the current prevalence of Helicobacter pylori infection. Gut, 35: 742-745.
PubMedDirect Link - Occhialini, A., A. Marais, M. Urdaci, R. Sierra, N. Muñoz, A. Covacci and F. Mégraud, 2001. Composition and gene expression of the cag pathogenicity island in Helicobacter pylori strains isolated from gastric carcinoma and gastritis patients in Costa Rica. Infect. Immun., 69: 1902-1908.
CrossRefPubMedDirect Link - Odenbreit, S., B. Gebert, J. Püls, W. Fischer and R. Haas, 2001. Interaction of Helicobacter pylori with professional phagocytes: Role of the cag pathogenicity Island and translocation, phosphorylation and processing of cagA. Cell. Microbiol., 3: 21-31.
CrossRefPubMedDirect Link - Odenbreit, S., J. Puls, B. Sedlmaier, E. Gerland, W. Fischer and R. Haas, 2000. Translocation of Helicobacter pylori cagA into gastric epithelial cells by type IV secretion. Science, 287: 1497-1500.
CrossRefPubMedDirect Link - Oliveira, A.G., A. Santos, J.B. Guerra, G.A. Rocha and A.M.C. Rocha et al., 2003. babA2- and cagA-positive Helicobacter pylori strains are associated with duodenal ulcer and gastric carcinoma in Brazil. J. Clin. Microbiol., 41: 3964-3966.
CrossRefPubMedDirect Link - Owen, R.J., T.M. Peters, R. Varea, E.L. Teare and S. Saverymuttu, 2001. Molecular epidemiology of Helicobacter pylori in England: prevalence of cag pathogenicity markers and IS605 prevalence in relation to patient age and severity of gastric disease. FEMS. Immunol. Med. Microbiol. 30: 65-71.
CrossRefPubMedDirect Link - Parsonnet, J., 2003. What is the Helicobacter pylori global reinfection rate? Can. J. Gastroenterol., 17: 46B-48B.
PubMedDirect Link - Proença Módena, J.L., A.I. Lopes Sales, G. Olszanski Acrani, R. Russo and M.A. Vilela Ribeiro et al., 2007. Association between Helicobacter pylori genotypes and gastric disorders in relation to the cag pathogenicity Island. Diagn. Microbiol. Infect. Dis., 59: 7-16.
CrossRefPubMedDirect Link - Pounder, R.E. and D. Ng, 1995. The prevalence of Helicobacter pylori in different countries. Aliment. Pharmacol. Ther., 9: 33-40.
PubMed - Salimkhani, E., R. Ranjbar, N. Sadeghifard, S. Morovvati, N. Jonaidi and M. Izadi, 2007. . An outbreak of gastroenteritis of unknown origin in Tehran, July 2003. Pak. J. Biol. Sci., 10: 1138-1140.
Direct Link - Ranjbar, R., N. Sadeghifard, A. Ahmadi, M. Izadi and J. Zaeimi-Yazdi et al., 2007. Antimicrobial susceptibility and AP-PCR typing of Acinetobacter spp. strains. Iran. J. Publ. Health, 36: 50-56.
Direct Link - Ranjbar, R., M.R. Pourshafie, N. Sadeghifard, A. Karami and M. Hamidian et al., 2008. Molecular characterization of epidemic isolates of Vibrio cholerae O1 by arbitrarily primed PCR (AP-PCR). Iran. J. Publ. Health, 37: 83-87.
Direct Link - Ranjbar, R., C. Mammina, M.R. Pourshafie and M.M. Soltan Dallal, 2008. Characterization of endemic Shigella boydii strains isolated in Iran by serotyping, antimicrobial resistance, plasmid profile, ribotyping and pulsed-field gel electrophoresis. BMC Res. Notes, 1: 74-74.
CrossRefPubMedDirect Link - Slater, E., R.J. Owen, M. Williams and R.E. Pounder, 1999. Conservation of the cag pathogenicity island of Helicobacter pylori and association with vacuolating cytotoxin alleles and IS605. Gastroenterology, 117: 1308-1315.
CrossRefPubMedDirect Link - Stein, M., R. Rappuoli and A. Covacci, 2000. Tyrosine phosphorylation of the Helicobacter pylori cagA antigen after cag-driven host cell translocation. Proc. Natl. Acad. Sci. USA., 97: 1263-1268.
PubMedDirect Link - Stone, G.G., D. Shortridge, R.K. Flamm, J. Beyer, A.T. Ghoneim and S.K. Tanaka, 1997. PCR-RFLP typing of ureC from Helicobacter pylori isolated from gastric biopsies during a European multi-country clinical trial. J. Antimicrob. Chemother., 40: 251-256.
PubMedDirect Link - Tomasini, M.L., S. Zanussi, M. Sozzi, R. Tedeschi, G. Basaglia and P. De Paoli, 2003. Heterogeneity of cag genotypes in Helicobacter pylori isolates from human biopsy specimens. J. Clin. Microbiol., 41: 976-980.
CrossRefPubMedDirect Link - Tan, H.J., A.M. Rizal, M.Y. Rosmadi and K.L. Goh, 2006. Role of Helicobacter pylori virulence factor and genotypes in non-ulcer dyspepsia. J. Gastroenterol. Hepatol., 21: 110-115.
CrossRefPubMedDirect Link - Yamazaki, S., A. Yamakawa, T. Okuda, M. Ohtani and H. Suto et al., 2005. Distinct diversity of vacA, cagA and cagE genes of Helicobacter pylori associated with peptic ulcer in Japan. J. Clin. Microbiol., 43: 3906-3916.
CrossRefPubMedDirect Link - Zhou, J., J. Zhang, C. Xu and L. He, 2004. Caga genotype and variants in Chinese Helicobacter pylori strains and relationship to gastroduodenal diseases. J. Med. Microbiol., 53: 231-235.
CrossRefPubMedDirect Link - Evans, D.G., D.M. Queiroz, E.N. Mendes and D.J. Evans Jr., 1998. Helicobacter pylori cagA status and s and m alleles of vacA in isolates from individuals with a variety of H. pylori-associated gastric diseases. J. Clin. Microbiol., 36: 3435-3437.
PubMed - Lin, C., S.C. Wu, S.C. Lee and K.S. Cheng, 2000. Genetic analysis and clinical evaluation of vacuolating cytotoxin gene A and cytotoxin-associated gene A in taiwanese Helicobacter pylori isolates from peptic ulcer patients. Scand. J. Infect. Dis., 32: 51-57.
PubMed - Smith, S.I., C. Kirsch, K.S. Oyedeji, A.O. Arigbabu and A.O. Coker et al., 2002. Prevalence of Helicobacter pylori vacA, cagA and iceA genotypes in Nigerian patients with duodenal ulcer disease. J. Med. Microbiol., 51: 851-854.
CrossRefPubMedDirect Link