
A. Mojtahedin
Physiology Division, Department of Basic Sciences, Faculty of Veterinary Medicine, Urmia University, P.O. Box 1177, Urmia 57135, Iran
E. Tamaddonfard
Physiology Division, Department of Basic Sciences, Faculty of Veterinary Medicine, Urmia University, P.O. Box 1177, Urmia 57135, Iran
A. Zanbouri
Physiology Division, Department of Basic Sciences, Faculty of Veterinary Medicine, Urmia University, P.O. Box 1177, Urmia 57135, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 22 | Page No.: 2573-2578







ABSTRACT
In this study, the effects of mepyramine (H1-receptor antagonist), famotidine (H2-receptor antagonist), physostigmine (a cholinesterase inhibitor) and atropine (muscarinic-receptor antagonist) have investigated on the formalin-induced nociception in rats. The effects of mepyramine and famotidine have also examined on nociceptive changes induced by physostigmine and atropine. Nociception was induced by intraplantar injection of formalin (50 μL, 1%) into the right hind paw and the time spent licking and biting of the injected paw, was taken as a measure of pain. Formalin induced a marked biphasic (first phase: 0-5 min and second phase: 15-45 min) pain response. The used drugs did not change the first phase of formalin-induced pain. Subcutaneous injection of physostigmine significantly (p<0.05) suppressed pain. Subcutaneous injection of atropine alone did not change the intensity of pain, but pretreatment with atropine significantly (p<0.05) prevented physostigmine-induced antinociception. Intraperitoneal injections of mepyramine and famotidine significantly (p<0.05) decreased pain response. Mepyramine did not significantly change, but famotidine significantly (p<0.05) prevented analgesic effect of physostigmine on pain. Atropine did not inhibit the antinociceptive effects of both mepyramine and famotidine on formalin-induced nociception. These results indicate that physostigmine through muscarinic cholinergic receptors suppresses the pain induced by formalin. Both H1 and H2 receptor antagonists produce antinociception. Histamine H2, but no H1 antagonists may be involved in physostigmine-induced antinociception.
PDF Abstract XML References Citation
How to cite this article
A. Mojtahedin, E. Tamaddonfard and A. Zanbouri, 2008. Effects of Mepyramine and Famotidine on the Physostigmine-Induced Antinociception in the Formalin Test in Rats. Pakistan Journal of Biological Sciences, 11: 2573-2578.
DOI: 10.3923/pjbs.2008.2573.2578
URL: https://scialert.net/abstract/?doi=pjbs.2008.2573.2578
DOI: 10.3923/pjbs.2008.2573.2578
URL: https://scialert.net/abstract/?doi=pjbs.2008.2573.2578
INTRODUCTION
Many antinociceptive tests in animals have developed for study the mechanisms of pain and analgesia (Le Bars et al., 2001). Formalin, as a nociceptive stimulus, has been frequently used to study of pain mechanisms in laboratory animals and according to these studies, a biphasic pattern of pain-related behaviors was produced by injection of small amounts (20-100 μL) of dilute solution (0.1-10%) of formalin into the various parts of body (Capone and Aloisi, 2004; Raboisson and Dallel, 2004). The first phase in turn may be attributed to a direct algogenic effect of formalin on the nociceptors and the second phase to release of local inflammatory mediators responsible for sensitization of primary and spinal sensory neurons and subsequent signal transduction into the brain (Hacimuftuoglu et al., 2006).
Acetylcholine plays pivotal roles in a diverse array of physiological processes such as learning, memory, anxiety, epilepsy, attention, cognition and consciousness (Hasselmo, 2006; File et al., 2000; Friedman et al., 2007; Wolf, 2006). Acetylcholine esterase controls the activity of acetylcholine in the sites of cholinergic transmission (Cummings, 2000). The role of acetylcholine, cholinergic agonists and cholinesterase inhibitors, collectively termed cholinomimetics, in the modulation of pain and analgesia has been established (Jones and Dunlop, 2007). Intra-hippocampal microinjection of acetylcholine and pilocarpine decreased frequency of discharge of pain-exited neurons and increased frequency of discharge of pain-inhibited neurons in the hippocampal CA1 in rats (Yang et al., 2008). Intrathecal injection of neostigmine, an acetylcholine esterase inhibitor, produced analgesia in a model of inflamed knee in rats (Buerkle et al., 1998). In tail flick and acetic acid-induced writhing tests in mice, antinociceptive effect of intraperitoneally injected physostigmine was reported by Patil and Kulkarni (1999).
Outside of the brain and in the peripheral tissues such as joints and skin, histamine stimulates nociceptive afferent fibers (Fu et al., 2005; Ting et al., 2007; Koppert et al., 2001). In the spinal cord level, intrathecal injection of histamine elicited a hyperalgesic response as assessed by tail flick test in mice (Sakurada et al., 2002). Within the brain, antinociceptive effects of the centrally administered histamine have been reported by Tamaddonfard and Rahimi (2004), Tamaddonfard et al. (2008a) and Mojtahedin et al. (2008). It is evident that several histamine H1, H2, H3 and H4 antagonists, but not all, produce antinociceptive effects in some animal models of pain. Antinociceptive effects of chorpheniramine (H1-receptor antagonist) and ranitidine (H2-receptor antagonist) in acetic acid-induced visceral pain was reported by Zanboori et al. (2008). Histamine H3 receptor agonist, imepip, attenuated formalin-induced pain and peripheral and central pretreatments with thioperamide (H3-receptor antagonist) reversed suppressive effect of imepip (Cannon et al., 2007). Histamine H4-receptor antagonists such as JNJ7777120 and VUF6002 has been reported to reduce hyperalgesia provoked by subplantar injection of carrageenan in rats (Coruzzi et al., 2007).
There are some interactions between histamine antagonists and cholinergic agents in brain and peripheral tissues. For example, in the physostigmine-induced yawning, involvement of both histamine H1 and H2 receptors has been reported by Tamaddonfard et al. (2008b). Moreover, histamine H1 receptors are involved in the neostigmine-induced central nervous system-mediated hyperglycemia (Nonogaki et al., 1993). It has been reported that histamine H2 receptor antagonists such as ranitidine, cimetidine and roxatidine do not interfere with muscarinic receptors in the contractile activity of fundus of stomach in guinea pigs (Milenov et al., 1995). H1-antihistaminics such as desloratadine and diphenhydramine, but not fexofenadine and cetirizine produced antimuscarinic effects on heart contractility (Liu et al., 2006).
The aim of the present study was to investigate the effects of mepyramine (H1-receptor antagonist), famotidine (H2-receptor antagonist), physostigmine (a cholinesterase inhibitor) and atropine (a muscarinic antagonist) on the formalin-induced pain response in rats. The interactions between histamine H1 and H2 antagonists with the effects of physostigmine and atropine on pain were also examined.
MATERIALS AND METHODS
Animals: Healthy adult male albino Wistar rats weighing 200-220 g were maintained in polypropylene cages with 6 rats in each cage with food and water available ad libitum, in a laboratory with controlled ambient temperature (20-23°C) and under a 12 h light-dark cycle (lights on 07:00 h). Six rats were used in each treatment. The experimental protocol was approved by the Laboratory Animal Care and Use Center of Urmia University. All experiments were conducted between 9:00 h and 15:00 h.
Drugs and treatments: Drugs used in the present study were mepyramine (pyrilamine), famotidine hydrochloride, physostigmine (eserin) and atropine sulfate. All drugs were purchased from Sigma-Aldrich Co., Steinheim, Germany. The drugs were dissolved in normal saline, expect of famotidine, which initially was dissolved in one drop of 1 M hydrochloric acid and then diluted with saline normal. Mepyramine at doses of 5, 10 and 20 mg kg-1 b.wt. and famotidine at doses of 10, 20 and 40 mg kg-1 b.wt., were intraperitoneally injected 30 min before induction of nociception. Physostigmine at doses of 0.05, 0.1 and 0.2 mg kg-1 b.wt. and atropine at a dose of 2 mg kg-1 b.wt., were subcutaneously administered 20 and 40 min before induction of formalin-induced pain, respectively. Atropine (2 mg kg-1 b.wt., s.c.) was injected 20 min before subcutaneous injection of physostigmine (0.1 mg kg-1 b.wt.). In combined treatments, intraperitoneal injections of mepyramine (10 mg kg-1 b.wt.) and famotidine (20 mg kg-1 b.wt.) were performed 10 min before physostigmine (0.1 mg kg-1 b.wt., s.c.) and 10 min after atropine (2 mg kg-1 b.wt., s.c.) administrations. Drug solutions were intraperitoneally injected in a volume of 1 mL kg-1 b.wt., using a 25-gauge injection needle. Subcutaneous injections of drug solutions were performed in a constant volume 0.2 mL per rat at the neck region using a 27-gauge injection needle.
Formalin test: Formalin test was used for induction of nociception. Before rats were pain tested, they were placed in a plexiglass observation chamber (30x30x25 cm) for 30 min on three successive days to minimize stress-activated pain suppressive mechanisms (Abbott and Bonder, 1997). The formalin test was applied as follows. Fifty microlitres of 1% formalin was injected subcutaneously into the ventral surface of right hind paw using a 29-gauge injection needle (Mojtahedin et al., 2008; Oritz and Castaneda-Hernandez, 2008). The rat was then placed in the observation chamber with a mirror mounted at 45° beneath the floor to allow an unobstructed view of the paw. The time spent licking and biting the injected paw was taken as a measure of nociceptive response and was recorded in five min intervals for 1 h. In the present study, data collected between 0 to 5 min post-formalin injection represented phase one (early phase) and data collected between 15-45 min after injection of formalin represented phase two (late phase). At the end of experiments, all rats were killed using high dose ether.
Statistical analysis: Data were expressed as Mean±SEM. Differences among treated groups were statistically evaluated using the one-way Analysis of Variance (ANOVA) followed by Duncan`s test. Differences were considered significant at p<0.05.
RESULTS AND DISCUSSION
Separate and combined treatments with mepyramine, famotidine, physostigmine and atropine produced no significant effects on the first phase of formalin-induced pain (Table 1, 2).
Subcutaneous injection of physostigmine at doses of 0.05, 0.1 and 0.2 mg kg-1 b.wt., significantly (0.05) suppressed late phase of pain. Atropine (2 mg kg-1 b.wt., s.c.), used alone, was without significant effect. Pretreatment with atropine (2 mg kg-1 b.wt., s.c.) before physostigmine (0.1 mg kg-1 b.wt., s.c.) prevented physostigmine-induced antinociception (Table 1).
Intraperitoneal injections of mepyramine at a dose of 5 mg kg-1 b.wt., did not produce any significant effect, whereas at doses of 10 and 20 mg kg-1 b.wt., mepyramine significantly (p<0.05) decreased second phase of nociceptive response. Intraperitoneal injections of famotidine at a dose of 10 mg kg-1 b.wt., produced no significant effect, whereas at doses of 20 and 40 mg kg-1 b.wt., famotidine significantly (p<0.05) decreased second phase of formalin-induce pain (Table 2).
Mepyramine (10 mg kg-1 b.wt., i.p.) before subcutaneous injection of physostigmine (0.1 mg kg-1 b.wt.) non-significantly inhibited the antinociceptive effect of physostigmine. Intraperitoneal injection of famotidine (20 mg kg-1 b.wt.) before physostigmine (0.1 mg kg-1 b.wt., s.c.) significantly (p<0.05) prevented analgesia induced by physostigmine. Atropine (2 mg kg-1 b.wt., s.c.) non-significantly reversed the effects of both mepyramine (10 mg kg-1 b.wt., i.p.) and famotidine (20 mg kg-1 b.wt., i.p.) on the formalin-induced pain (Table 3).
The results presented here showed that physostigmine produced antinociception and atropine prevented physostigmine-induced analgesia. This indicates that physostigmine through muscarinic cholinergic receptors may be involved in pain modulation. Physostigmine is a major alkaloid found in the seeds of the fabaceous plant Physostigma venenosum and is a powerful and reversible acetylcholine esterase inhibitor that effectively increases concentration of acetylcholine in the sites of cholinergic transmission (Zhao et al., 2004). Several lines of evidence suggest that physostigmine influences modulation of pain. Subcutaneous injection of physostigmine produced antinociceptive effect in spinal nerve ligation model of neuropathic pain in rats. Atropine (muscarinic receptor antagonist) but not mecamylamine (nicotinic receptor antagonist) reversed antinociceptive effect of physostigmine (Poyhia et al., 1999). Moreover, in tail flick and acetic acid-induced writhing tests in mice, antinociceptive effect of intraperitoneally injected physostigmine was reported by Patil and Kulkarni (1999). Only in one study, suppressive effect of intrathecally-administered physostigmine on both phases of formalin-induced pain was reported in rats (Yoon et al., 2003).
| Table 1: | Effects of physostigmine and atropine on the durations(s) of licking and biting of the injected paw after intraplantar injection of formalin in rats |
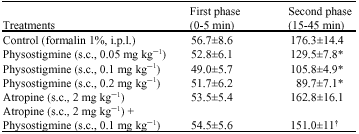 | |
| Values are Mean±SEM, n = 6 in each group, *p<0.05 vs. normal saline groups, xp<0.05 vs. physostigmine (0.1 mg kg-1) group, one way ANOVA followed by Duncan`s Test, i.p.l.: Intraplantar, s.c.: Subcutaneous | |
| Table 2: | Effects of mepyramine and famotidine on the durations(s) of licking and biting of the injected paw after intraplantar injection of formalin in rats |
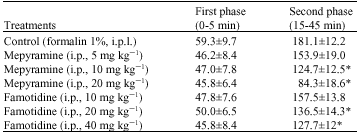 | |
| Values are Mean±SEM, n = 6 in each group, *p<0.05 vs. normal saline groups, one way ANOVA followed by Duncan`s Test, i.p.l.: Intraplantar, i.p.: Intraperitoneal | |
| Table 3: | Effects of mepyramine and famotidine on the changes induced by physostigmine and atropine in the durations(s) of licking and biting of the injected paw after intraplantar injection of formalin in rats |
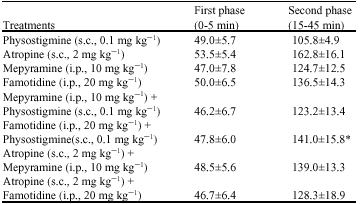 | |
| Values are Mean±SEM, n = 6 in each group, *p<0.05 vs. physostigmine (s.c., 0.1 mg kg-1) group, one way ANOVA followed by Duncan`s Test, s.c.: Subcutaneous, i.p.: Intraperitoneal | |
Neostigmine, a cholinesterase inhibitor, suppressed nociception in knee joint model and physostigmine attenuated pain in spinal nerve ligature model of neuropathic pain. In addition, atropine blocked neostigmine- and physostigmine-induced antinociception in both joint and neuropathic models of pain, suggesting a role for muscarinic cholinergic receptors in modulating cholinesterase inhibitors-induced antinociception (Buerkle et al., 1998; Poyhia et al., 1999).
In then present study, both mepyramine and famotidine produced antinociception. This indicates that both histamine H1 and H2 antagonists may have analgesic properties. It has been reported that some of histamine H1 and H2 antagonists produce antinociception in laboratory animal models of nociceptive tests (Raffa, 2001). In formalin test in mice, subcutaneous injections of dexchlorpheniramine (H1-receptor antagonist) and ranitidine (H2-receptor antagonist) suppressed formalin-induced nociceptive behavior (Farzin and Nosrati, 2007). Moreover, in p-benzoquinine- and acetic acid- writhing tests in mice, antinociceptive effects of mepyramine, ReN 1869 (a selective histamine H1 receptor antagonist) and famotidine were reported by Abacioglu et al. (1993), Olsen et al. (2002) and Girard et al. (2004). Intraperitoneal injections of chlorpheniramine and ranitidine produced antinociception in the acetic acid-induced visceral nociception in rats (Zanboori et al., 2008).
In the present study, famotidine but not mepyramine inhibited physostigmine-induced analgesia and atropine did not reverse the suppressive effects of both mepyramine and famotidine. These results indicate that there may be an interaction between histamine H2 but not H1 antagonists and physostigmine in the modulation of pain and analgesia. In guinea pig trachealis muscle preparations, mepyramine in comparison with H1-receptor antagonists such as cyproheptadine, promethazine, desloratadine, loratadine, chlorpheniramine showed a minimal anticholinergic activity (Orzechowski et al., 2005). Moreover, in the hippocampus of anaesthetized rats, intraperitoneal injection of mepyramine did not change the increase in acetylcholine release evoked by electrical stimulation of the tuberomammillary nucleus (Mochizuki et al., 1994). In addition, diphenhydramine but not mepyramine suppressed tacrine (a cholinomimetic drug)-induced tremulous jaw movements in rats (Carlson et al., 2000). It has been found that ranitidine but not famotidine increases both resting and electrical stimulation-evoked release of acetylcholine in the guinea pig myenteric plexus (Poli et al., 1990). In addition, both nizatidine (H2-receptor antagonist) and ranitidine but not famotidine produced stimulatory effects on both acetylcholinesterase activity and HCO–3 secretion in the duodenum in rats (Mimaki et al., 2001). Moreover, cimetidine completely but famotidine slightly inhibited the acid secretion evoked by vagal stimulation, bethanechol and histamine in the isolated mouse whole stomach (Watanabe et al., 1993). The differences between findings may be associated to the kind of experiment applied and to the kind of histamine antagonists used. However, there is not any report identifying the interaction between H1 and H2 antagonists with physostigmine in modulation of pain. In the other physiological functions such as memory and learning, yawning, gastric acid secretion, existence of negative and positive interactions have been reported by Eidi et al. (2003), Tamaddonfard et al. (2008) and Watanabe et al. (1993).
In conclusion, the results presented here indicate that physostigmine through muscarinic receptors is able to suppress the inflammatory pain induced by formalin. Both H1 and H2 receptor antagonists produce antinociception by reducing the intensity of pain in late phase. Histamine H2, but not H1 antagonists may be involved in physostigmine-induced antinociception.
REFERENCES
- Abacioglu, N., A. Bediz, L. Cakici, B. Tuncutan and I. Kanzik, 1993. Antinociceptive effect of H1-and H2-antihistaminics in mice. Gen. Pharmacol., 24: 1173-1176.
PubMed - Abbott, F.V. and M. Bonder, 1997. Options for management of acute pain in the rat. Vet. Rec., 140: 553-557.
PubMed - Buerkle, H., M. Boschin, M.A.E. Marcus, G. Brodner, R. Wutsen and H. Van Aken, 1998. Central and peripheral analgesia mediated by the acetylcholinesterase-inhibitor neostigmine in the rat inflamed knee joint model. Anesth. Analg., 86: 1027-1032.
PubMed - Carlson, B.B., J.T. Trevitt and J.D. Salamone, 2000. ffects of H1 antagonists on cholinomimetic-induced tremulous jaw movements: Studies of diphenhydramine, doxepin and mepyramine. Pharmacol. Biochem. Behav., 65: 683-689.
PubMed - Cannon, K.F., R. Leurs and L.B. Hough, 2007. Activation of peripheral and spinal histamine H3 receptors inhibits formalin-induced inflammation and nociception, respectively. Pharmacol. Biochem. Behav., 88: 122-129.
PubMed - Capone, F. and A.M. Aloisi, 2004. Refinement of pain evaluation techniques. The formalin test. Ann. Ist. Super Sanita., 40: 223-229.
PubMed - Cummings, J.L., 2000. Cholinesterase inhibitors: A new class of psychotropic compounds. Am. J. Psychiatry, 157: 4-15.
PubMed - Eidi, M., M.R. Zarrindast, A. Eidi, S. Oryan and K. Parivar, 2003. Effects of histamine and cholinergic systems on memory retention of passive avoidance learning in rats. Eur. J. Pharmacol., 465: 91-96.
PubMed - Farzin, D. and F. Nosrati, 2007. Modification of formalin-induced nociception by different histamine receptor agonists and antagonists. Eur. Neuropsychopharmacol., 17: 122-128.
PubMed - File, S.E., P.J. Kenny and S. Cheeta, 2000. The role of dorsal hippocampal serotonergic and cholinergic systems in the modulation of anxiety. Pharmacol. Biochem. Behav., 66: 65-72.
PubMed - Friedman, A., C.J. Behrens and U. Heineman, 2007. Cholinergic dysfunction in temporal lobe epilepsy. Epilepsia, 48: 126-130.
PubMed - Girard, P., Y. Pansart, M.C. Coppe, D. Verniers and J.M. Gillardin, 2004. Role of histaminergic system in nefopam-induced antinociception in mice. Eur. J. Pharmacol., 503: 63-69.
PubMedDirect Link - Hacimuftuoglu, A., C.R. Handy, V.M. Goettl, C.G. Lin, S. Dane and R.L. Jr. Stephens, 2006. Antioxidants attenuate multiple phases of formalin-induced nociceptive response in mice. Behav. Brain Res., 173: 211-216.
PubMed - Hasselmo, M.E., 2006. The role of acetylcholine in learning and memory. Curr. Opin. Neurobiol., 16: 710-715.
PubMed - Jones, P.G. and J. Dunlop, 2007. Targeting the cholinergic system as a therapeutic strategy for the treatment of pain. Neuropharmacology, 53: 197-206.
PubMed - Koppert, W., P. Martus and P.W. Reeh, 2001. Interactions of histamine and bradykinin on polymodal C-fibers in isolated rat skin. Eur. J. Pain, 5: 97-106.
PubMed - Le Bars, D.L., M. Gozariu and S.W. Cadden, 2001. Animal models of nociception. Pharmacol. Rev., 53: 597-652.
PubMedDirect Link - Liu, H., Q. Zheng and J.M. Farley, 2006. Antimuscarinic actions of antihistamines on the heart. J. Biomed. Sci., 13: 395-401.
PubMed - Milenov, K., S. Todorov, M. Vassileva, R. Zamfirova and A. Shahbazian, 1995. Different effects of H1 and H2 blockers on the tone and the contractile activity of guinea pig stomach fundus. Methods Find Exp. Clin. Pharmacol., 17: 609-613.
PubMed - Mimaki, H., S. Kawauchi, S. Kagawa, S. Ueki and K. Takeuchi, 2001. Bicarbonate stimulatory action of nizatidine a histamine H2-receptor antagonist, in rat duodenums. J. Physiol. (Paris), 95: 165-171.
PubMed - Mochizuki, T., K. Okakura-Mochizuki, A. Horii, Y. Yamamoto and A. Yamatodani, 1994. Histaminergic modulation of hippocampal acetylcholine release in vivo. J. Neurochem., 62: 2275-2282.
PubMed - Nonogaki, K., X. Li, T. Tamagawa, G. Watanabe and Y. Hiyoshi et al., 1993. Role of brain histamine H1- and H2-receptors in neostigmine-induced hyperglycemia in rats. Life Sci., 52: PL107-PL110.
PubMed - Olsen, U.B., C.T. Eltrop, B.K. Ingvargsen, T.K. Jorgensen and J.A. Lundbaek et al., 2002. ReN 1869 a novel tricyclic antihistamine, is active against neurogenic pain and inflammation. Eur. J. Pharmacol., 435: 43-57.
PubMed - Oritz, M.I. and G. Castaneda-Hernandez, 2008. Examination of the interaction between peripheral lumiracoxib and opioids on the 1% formalin test in rats. Eur. J. Pain, 12: 233-241.
PubMed - Orzechowski, R.F., D.S. Currie and C.A. Valancius, 2005. Comparative anticholinergic activities of 10 histamine H1 receptor antagonists in two functional models. Eur. J. Pharmacol., 506: 257-264.
PubMed - Patil, C.S. and S.K. Kulkarni, 1999. The morphine sparing effect of physostigmine. Methods Find Exp. Clin. Pharmacol., 21: 523-527.
PubMed - Poli, E., G. Coruzzi and G. Bertaccini, 1990. Ranitidine but not famotidine releases acethylcholine from the guinea pig myenteric plexus. Agents Actions, 30: 191-194.
PubMed - Poyhia, R., V.K. Kontinen, S. Paananen and E. Kalso, 1999. Systemic physostigmine shows antiallodynic effect in neuropathic rats. Anesth. Analg., 89: 428-433.
PubMed - Sakurada, S., T. Orito, C. Sakurada, T. Sato, T. Hayashi and J.I. Mobarakeh et al., 2002. Possible involvement of tachykinin NK1 and NMDA receptors in histamine-induced hyperalgesia in mice. Eur. J. Pharmacol., 434: 29-34.
PubMed - Tamaddonfard, E. and S. Rahimi, 2004. Central effect of histamine and peripheral effect of histidine on the formalin-induced pain response in mice. Clin. Exp. Pharmacol. Physiol., 31: 518-522.
PubMed - Tamaddonfard, E., E. Khalilzadeh, N. Hamzeh-Gooshchi and S. Seiednejhad-Yamchi, 2008. Central effect of histamine in a rat model of acute trigeminal pain. Pharmacol. Rep., 60: 219-224.
PubMed - Ting, E., R.C. Roveroni, L.F. Ferrari, C.M. Lotufo and M.C. Veiga et al., 2007. Indirect mechanism of histamine-induced nociception in temporomandibular joint of rats. Life Sci., 81: 765-771.
PubMed - Watanabe, K., S. Yano, M. Yamamoto and S. Kanaoka, 1993. Comparative effects of cimetidine and famotidine on the vagally stimulated acid secretion in the isolated mouse whole stomach. Jpn. J. Pharmacol., 61: 229-236.
PubMed - Yang, X.F. Y. Xiao and M.Y. Xu, 2008. Both endogenous and exogenous ACh plays antinociceptive role in the hippocampus CA1 of rats. J. Neural Transm., 115: 1-6.
PubMed - Yoon, M.H., J.I. Choi and S.W. Jeong, 2003. Antinociception of intrathecal cholinesterase inhibitors and cholinergic receptors in rats. Acta Anaesthesiol. Scand., 47: 1079-1084.
PubMed - Zanboori, A., E. Tamaddonfard and A. Mojtahedin, 2008. Effects of chlorpheniramine and ranitidine on the visceral nociception induced by acetic acid in rats: Role of opioid system. Pak. J. Biol. Sci., 11: 2428-2432.
CrossRefPubMedDirect Link - Zhao, B.M., S.M. Moochhala and S.Y. Tham, 2004. Biologically active components of Physostigma venenosum. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci., 812: 183-192.
PubMed