
Duangporn Kantachote
Department of Microbiology, Faculty of Science, Prince of Songkla University, Hat-Yai 90112, Thailand
Wilawan Charernjiratrakul
Department of Microbiology, Faculty of Science, Prince of Songkla University, Hat-Yai 90112, Thailand
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 22 | Page No.: 2545-2552







ABSTRACT
Lactic Acid Bacteria (LAB) from fermented plant beverages were selected based on their antibacterial actions against potential food borne pathogenic bacteria (Staphylococcus aureus PSSCMI 0004, Escherichia coli PSSCMI 0001, Salmonella typhimurium PSSCMI 0034 and Vibrio parahaemolyticus VP 4). Antibacterial activities were measured using an agar spot method. The Lactobacillus plantarum W90A strain isolated from a wild forest noni (Morinda coreia Ham) beverage was used as an inoculant. Three different inoculation procedures were conducted with the fruit of wild forest noni fermentations to establish which one was the best for controlling the numbers of yeast in the finished product. A 5% inoculum of L. plantarum W90A (LAB set), initial cell density 8.6 log cfu mL-1, produced a better product and inhibitory properties against the test organisms, particularly E. coli PSSCMI 0001 than one with no inoculum or with a 5% inoculum from a previous natural fermented product. An LAB inoculum resulted in a reduced total bacterial count and no yeast throughout fermentation period (90 days). The lower yeast resulted in a reduction of the ethanol content to 2.9 g L-1 compared to 12.2 g L-1 in the culture with no inoculum. The highest acidity (1.3-1.4%) with the same pH (3.3) was observed in both sets of inoculated fermentations, whereas the uninoculated set gave a pH value of 3.7 (1.2% acidity).
PDF Abstract XML References Citation
How to cite this article
Duangporn Kantachote and Wilawan Charernjiratrakul, 2008. Selection of Lactic Acid Bacteria from Fermented Plant Beverages to Use as Inoculants for Improving the Quality of the Finished Product. Pakistan Journal of Biological Sciences, 11: 2545-2552.
DOI: 10.3923/pjbs.2008.2545.2552
URL: https://scialert.net/abstract/?doi=pjbs.2008.2545.2552
DOI: 10.3923/pjbs.2008.2545.2552
URL: https://scialert.net/abstract/?doi=pjbs.2008.2545.2552
INTRODUCTION
There have been many reports that Lactic Acid Bacteria (LAB) exert strong antagonistic activity against food spoilage and pathogenic bacteria, particularly those involved with food borne diseases (Ivanova et al., 2000; Sjogren et al., 2003; Chung and Yousef, 2005). Some products of lactic acid fermentation such as, organic acids, diacetyl, ethanol, hydrogen peroxide, and bacteriocins are well known preservatives that possess antibacterial activity and may help to control pathogenic bacteria (Lindgren and Dobrogosz, 1990; Kang and Fung, 1999; Chung and Yousef, 2005). Organic acids such as lactic acid produced by LAB strongly inhibit some enteropathogens (Fooks and Gibson, 2002). Zalan et al. (2005) demonstrated that Lactobacillus species (4 isolates) grown in MRS broth produced from 2-6 μg mL-1 hydrogen peroxide after 72 h of incubation and this was much higher than that produced in tomato juice broth.
In Thailand, Fermented Plant Beverages (FPBs) produced by lactic acid fermentation are consumed by many Thai people who believe that these products have a high nutrient value and also have antimicrobial activities. The antimicrobial activities in the FPBs may be produced by LAB but others may be extracted from the plant materials used in the process (Kantachote et al., 2005a). Thailand is located in the tropical zone, and diarrheal diseases caused by food-borne infections and food toxicity is endemic (FAO/WHO, 2002) and locally produced FPBs are thought by some Thai people to be able to protect themselves from these food borne illnesses. Although this might be true, there are some major problems involved in scaling up the production of these local FPBs to allow them to be produced as a commercially viable product. One such problem is the high contamination by yeasts in their finished products (Prachyakij et al., 2007) due mostly to the fact that the finished product is not pasteurized. That means that the shelf life of the finished product is short and there is no certainty that potential spoilage and pathogenic microbes have been significantly reduced. Consumers and producers also believe that heat may destroy nutrients and other valuable compounds such as anti-oxidants, so tend to avoid procedures that may involve heating that could reduce microbial contamination. To help local communities to improve and control the quality of their finished FPBs in a so called natural way the use of standard characterized inoculants might help. The aim of this study was, therefore to screen LAB for FPBs that can inhibit potential food borne pathogenic bacteria and to examine their use as starter cultures to improve the quality of the fermented plant beverage.
MATERIALS AND METHODS
Lactic acid bacteria: Sixty LAB were previously isolated from 2 fermented plant beverages, one produced from wild forest noni and the other from phomnang seaweed (Gracilaria fisheri) (Kantachote and Charernjiratrakul, 2007). Forty of these isolates that grew rapidly on MRS agar, were chosen to investigate their antibacterial activity against bacterial food borne pathogens.
Indicator bacteria: The following organisms Staphylococcus aureus PSSCMI 0004, Escherichia coli PSSCMI 0001, Salmonella typhimurium PSSCMI 0034 and Vibrio parahaemolyticus VP 4 were used as bacterial indicators to evaluate the antibacterial effects of LAB isolated from the FPBs. The test strains isolated from clinical specimens during 2004-2005 were obtained from the Department of Microbiology, Faculty of Science, Prince of Songkla University, Thailand.
Selection steps of LAB: Three steps were used to screen for LAB isolates with antibacterial activities using the agar spot method of Spelhaug and Halander (1989) and Schillinger and Lucke (1989). Briefly, cultures were first grown with MRS broth, containing 2% glucose for 18 h and 5 μL of each isolate (107 cells mL-1) was spotted onto the surface of the MRS agar plate (4 isolates/plate). Each indicator strain (106 cells mL-1) was inoculated into 7 mL of soft BHI (Brain Heart Infusion) agar (0.7% agar) and poured over the plate on which the producers (LAB) had grown. There were four replicates for each LAB isolate. For the first step, all plates were incubated at 35°C for 24 h and no attempt was made to control the production of either hydrogen peroxide or acids. The plates were checked for inhibition zones and the inhibition was scored to 4 levels as follows: <5-5 mm = +, >5-10 mm = ++, >10-15 mm = +++, >15 mm = ++++.
To limit hydrogen peroxide production, a similar procedure to the above process was used but adapted by placing the plates in an anaerobic jar for incubation. Inhibition zones were ranked as follows: >4-5 mm = +, >5-6 mm = ++, >6-7 mm = +++, >7-8 mm = ++++, >8-9 mm = +++++, >9 mm = ++++++.
To limit productions of both organic acids and hydrogen peroxide MRS plates with only 0.2% glucose were used with incubation again in anaerobic jars. Inhibition zones were classified as follows: >2-3 mm = +, >3-4 mm = ++, >4-5 mm = +++, >5-6 mm = ++++.
Only results of the LAB isolates that gave good inhibition zones against the bacterial indicator strains have been presented as diameters of their zones of inhibition.
Wild forest noni fermentation: The wild forest noni tree (Morinda coreia Ham) is a medicinal plant commonly found in all parts of Thailand. In earlier study we had observed that fermented beverages produced from the fruit of this plant did inhibit the growth of potential enteropathogenic (Kantachote et al., 2005b). Ripe fruit of wild forest noni often has a bad smell and using a ripe fruit would also have introduced another difficult to control variable i.e., the possibility of adding more potential spoilage organisms to the fermentation. Our previous work we therefore chose to use non ripened fruit and L. plantarum W90A was one of isolate from this beverage (Kantachote and Charernjiratrakul, 2007). However, now we investigated to use inoculants of a selected strain from this study, L. plantarum W90A, or a previous fermented beverage that might be useful to allow better quality control with a fermentation starting with ripened noni. L. plantarum W90A was prepared to use as starter culture by using cells in a late log phase from MRS medium. Cells were separated by centrifugation (6000 rpm, 15 min) and washed twice by normal saline solution. Finally, cells pellet was resuspended by sterile distilled water to obtain approximate x1010 cfu mL-1 cell density based on preliminary test using pour plate method. Three different inoculums were used to inoculate and initiate the fermentation of the fruit of wild forest noni as follows: 5% inoculum of a previously fermented wild forest noni preparation (FWB set), a 5% of L. plantarum W90A culture (LAB set) and no additional inoculum as a control set. Each 28 liter fermentation bucket contained 6 kg of ripe fruit, 2 kg of sugarcane and 20 L of tap water while the space above the fermentation liquid was filled with a water filled plastic bag to make the fermentation virtually anaerobic (Kantachote and Charernjiratrakul, 2007). Three replicates were carried out for each treatment and sampled at days 0, 1, 2, 3, 4, 5, 6, 7, 14, 21, 30, 45, 60, 75 and 90 in order to monitor the characteristics of the fermented beverage and any microbiological succession.
Microbial enumeration: Microbial counts were made at every sampling time using standard methods (FDA, 2001). Ten fold dilutions starting with 25 mL of each sample added to 225 mL normal saline solution to obtain a 10-1 dilution and then appropriate dilutions were used for pour plate counting of LAB and Total Bacterial Count (TBC). De Man Rogosa Shape (MRS) medium was used for the LAB, whereas Plate Count Agar (PCA) was used for TBC. Potato Dextrose Agar (PDA) was used for counting molds and yeasts by the spread plate technique. All plates were incubated at room temperature (28±3°C).
Determination of the chemical properties of fermented beverages: Standard analytical methods (AOAC, 2002) were used to determine pH, titratable acidity and Total Acidity (TA) was calculated in the form of lactic acid. Total Sugar (TS) in the form of glucose was examined using the phenol sulfuric method (Dubois et al., 1956). All chemical properties were monitored at every sampling time, except for the organic acids (lactic acid and acetic acid) and alcohols (ethanol and methanol) that were only measured at days 0, 30, 60 and 90. Those compounds were estimated by gas chromatography as described by Yang and Choong (2001).
Investigation of antibacterial activity of fermented beverage: Using the same set of test organisms used for the LAB screening a cup well diffusion method was also used to test for the antagonistic activity of the fermented beverages. Following at least two subcultures (24 h, 37°C) on fresh (TSA: tryptic soy agar or TSA plus 2%NaCl for V. parahaemolyticus VP 4) plates prior to the experiment. Each actively growing culture from the plate was adjusted to 0.5 (McFarland standard) using 0.85% NaCl and then swabbed over the surface of a TSA plate. The fermented beverage samples, collected at different fermentation times (0, 30, 60 and 90 days) were filter sterilized using a 0.45 μm filter and 125 μL of each filtered sample of fermented wild forest noni beverage was added into each cup well.
RESULTS
Selection of LAB isolates based on their antibacterial activities: LAB isolates growing aerobically can produce both organic acids and hydrogen peroxide as metabolites (Hammes and Vogel, 1995; Adams and Moss, 2000). Eighteen LAB isolates (45%) produced good inhibition zones against all test organisms and those producing inhibition zones >10-15 mm i.e., +++ and >15 mm, ++++ were selected for further studies. This involved only four strains P45B, P75B, W60A and W90A (Table 1). The order of sensitivity of the test bacteria to the LAB isolates (40 isolates) grown with aerobic conditions was V. parahaemolyticus VP 4 > S. typhimurium PSSCMI 0034 > S. aureus PSSCMI 0004 > E. coli PSSCMI 0001 (data not shown).
When the 4 selected LAB isolates were grown with MRS under anaerobic conditions so that production of hydrogen peroxide was limited and the order of sensitivity was, V. parahaemolyticus VP 4 with an inhibition zone of 9-10 mm followed by S. typhimurium PSSCMI 0034 (inhibition zone: 7-8 mm), and S. aureus (inhibition zones: 7-8 mm). E. coli was the most resistant with an inhibition zone of between 5-6 mm. Strains P75B and W90A were better inhibitors of all test organisms than the other strains and both strains had been previously identified as Lactobacillus plantarum (Kantachote and Charernjiratrakul, 2007).
When production of both organic acids and hydrogen peroxide was limited by anaerobic growth and reduced (0.2%) glucose only a few of the selected LAB isolates exhibited good antibacterial actions against all test organisms (Table 1). This indicates that these isolates must produce antibacterial compounds other than hydrogen peroxide and organic acids and were therefore further investigated. Strain W90A, isolated from fermented wild forest noni, was the most effective strain to inhibit all test organisms (inhibition zones: 5-6 mm). Another selected strain P75B, isolated from the phomnang seaweed beverage, gave a similar inhibition zone against bacterial indicators (inhibition zone: 3-6 mm); however, strain W90A gave a stronger inhibition of E. coli PSSCMI 0001 than the strain P75B. Hence, isolate W90A was selected for investigating its potential for use as a starter cultures.
Effects of different inoculation procedures on wild forest noni fermentation: All fermentations started with the same initial pH of 4.5 and after day 1 dropped to 4.0; however, at day 90 the control set i.e. with no added inoculum had a higher pH (3.7) than that found in the other 2 sets (pH 3.3) (data not shown). The TA levels were in agreement with the pH values being lowest (1.2%) in the control and in other sets were 1.3% (FWB) and 1.4% (LAB) (Fig. 1). The acidity levels were also correlated with the amounts of sugar used as the initial TS (13%) decreased to 4.65% in the control and 4.15-3.83% in the FWB and LAB, respectively (Fig. 1).
The numbers of TBC and LAB, including their changed numbers, were similar throughout all sets of wild forest noni beverage fermentations. This meant that most of the TBC cells were LAB cells. Therefore only the numbers of LAB and yeast are shown (Fig. 2). Yeast was not detected throughout the fermentation in any set inoculated with LAB. The initial yeast count in the FWB set was higher than that in the control set; however, after day 14 no yeast was detected in the FWB set. In contrast, at the end of the fermentation on day 90 the control set had 1.6 log cfu mL-1 yeast. Initial counts of LAB was 8.6 log cfu mL-1 in the LAB set, 4.6 log cfu mL-1 in the FWB set and 2.8 log cfu mL-1 in the control set. After day 1 of fermentation the LAB counts dropped about 3 log cycles in the LAB set then LAB increased and reached a maximum of 8.2 cfu mL-1 at day 5. Similar changes of LAB counts occurred in all 3 procedures, a significant drop after day 1 followed by an increase to reach a maximum at day 5 in the LAB set or day 6 in the FWB and control sets at 7.9 cfu mL-1, following day 5 or 6 LAB counts declined to 2.5 cfu mL-1 at day 90 in all sets of fermentation.
| Table 1: | Inhibitory effect of isolated lactic acid bacteria against food borne pathogenic bacteria under various culture conditions |
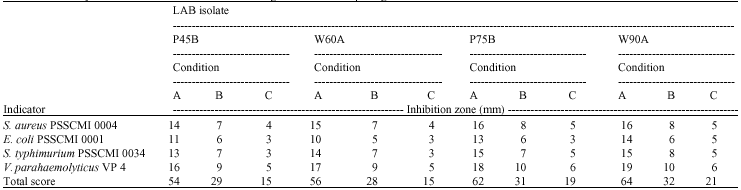 | |
| A = Combination of organic acids and hydrogen peroxide production, B = Unlimited production of organic acids but limited hydrogen peroxide production, C = Limitation of both organic acids and hydrogen peroxide production | |
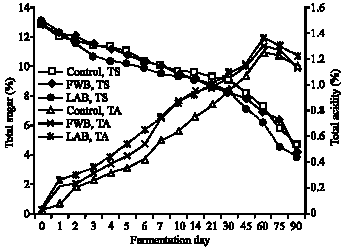 | |
| Fig. 1: | Effects of different inoculation procedures on changes of total sugar and total acidity in a wild forest noni fermentation |
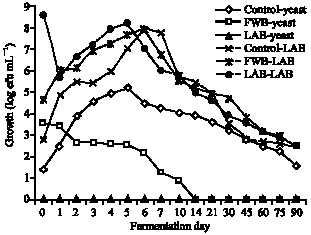 | |
| Fig. 2: | Effects of different inoculation procedures on populations of lactic acid bacteria and yeast in a wild forest noni fermentation |
Effects of different inoculation procedures on organic acids and alcohol levels in the fermented beverages relative to their ability to inhibit test organisms: After day 60 in all sets of fermentation, the amount of acetic acid (6.5-7.5 g L-1) was higher than that of lactic acid (5.0- 5.3 g L-1) (Table 2). However, at day 90 the amount of acetic acid in all sets significantly decreased, whereas the amount of lactic acid decreased only slightly (Table 2). The highest amount of acetic acid was found in the LAB set, while similar amounts were detected in both the control and FWB sets. In contrast, at the end of fermentation the control set had the highest level of ethanol (12.2 g L-1) followed by the LAB set (8.6 g L-1) and the lowest level in the FWB set (2.9 g L-1). The amount of methanol was less than 0.20 g L-1 in all sets throughout the fermentation.
No antibacterial activity was observed at day 0 in any set of wild forest noni fermentations but increased with the time of fermentation. The best inhibition was obtained at days 60 and 90 (Table 3). A similar result to control most of test organisms was observed between LAB and FWB sets and it was higher than that found in the control set. However, inhibitory effect against E. coli PSSCMI 0001 of beverages was in the order of LAB set > FWB set > control set and the order of the test strains sensitivity was V. parahaemolyticus VP4 > S. aureus PSSCMI 0004 > S. typhimurium PSSCMI 0034 > E. coli PSSCMI 0001. Atday 60 the beverage from the LAB set produced an inhibition zone with E. coli PSSCMI 0001 of 12.4 mm while the other 2 sets produced their best inhibition at day 90 with a 9.20 mm inhibition zone for the control and a 12.2 mm zone for the FWB set.
| Table 2: | Effects of different inoculation procedures on organic acids and alcohols contents in a wild forest noni fermentation |
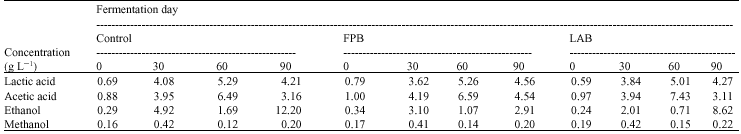 | |
| FPB = 5% inoculum of a previously fermented wild forest noni, LAB = 5% of Lactobacillus plantarum W90A starter day 60 the beverage from the LAB set produced an inhibition zone with E. coli PSSCMI 0001 of 12.4 mm while the other 2 sets produced their best inhibition at day 90 with a 9.20 mm inhibition zone for the control and a 12.2 mm zone for the FWB set. | |
| Table 3: | Effects of different inoculation procedures on the inhibition of food borne pathogenic bacteria in a wild forest noni fermentation |
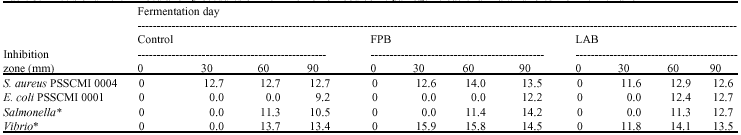 | |
| *: Salmonella typhimurium PSSCMI 0034, V. parahaemolyticus VP 4, FPB = 5% inoculum of a previously fermented wild forest noni, LAB = 5% of Lactobacillus plantarum W90A starter | |
DISCUSSION
Selection of lactic acid bacteria to use as inoculants: An agar spot test was used to demonstrate antibacterial activities this indicates that the inhibitory metabolites produced by the LAB isolates were extracellular and diffusible. LAB isolates incubated under aerobic conditions gave larger inhibition zones against the test organisms than when grown with anaerobic conditions (Table 1). Aerobically LAB produce more organic acids a larger reduction in the pH and also produce hydrogen peroxide (Lindgren and Dobrogosz, 1990; Adams and Moss, 2000). It has long been recognized that hydrogen peroxide as an oxidizing agent can inhibit both Gram negative and positive bacteria (Yap and Gilliland, 2000; Zalan et al., 2005). Therefore when production of hydrogen peroxide is inhibited by growth in anaerobic conditions, most LAB isolates had reduced levels of inhibition but not as much as when the level of acid production was further reduced in the control by a reduction of the glucose substrate.
In most situations S. aureus PSSCMI 0004 was more sensitive than that observed in S. typhimurium PSSCMI 0034 and E. coli PSSCMI 0001 was the most resistant. This means that there may be antibacterial metabolites apart from the organic acids and hydrogen peroxide that inhibited S. aureus PSSCMI 0004. Bacteriocins for example are more effective against Gram positive bacteria than Gram negative bacteria (Hernnadez et al., 2005; Osmanagagaoglu et al., 2005). Therefore, the possibility that bacteriocins may be produced by the selected LAB isolates will be investigated in a further study. Gram negative bacteria are generally not sensitive to bacteriocins from LAB unless the barrier function of their outer membrane is disrupted by treatment with chelating agents such as EDTA (Adams and Moss, 2000). Clavero and Beuchat (1996) have also reported that E. coli O157: H7 is more tolerant to some organic acids than many other infectious pathogens and can survive well in acidic foods and beverages.
The resistance of Gram negative bacteria towards antibacterial substances is related to their lipopolysaccharides outer membrane (Gao et al., 1999). Results indicate that V. parahaemolyticus VP 4 and S. typhimurium PSSCMI 0034 were very sensitive to organic acids in the absence of hydrogen peroxide production (Table 1). In contrast, the effect of organic acids on E. coli PSSCMI 0001 was less obvious. Perhaps the outer membrane of E. coli may differ from other Gram negative bacteria such as V. parahaemolyticus and S. typhimurium. Similar results have been reported by Simango and Rukure (1992) as pathogenic E. coli strains were more tolerant to the low pH of the fermented foods than Salmonella. In addition, there are reports that some Gram negative bacteria have an acid-tolerant response system that protects them against severe acid stress for longer periods (Foster, 1991).
It is not surprising that both the LAB strains selected from those isolated from Thai fermented plant beverages are strains of Lactobacillus plantarum as this organism is normally found in fermented plants and vegetables (Daeschel et al., 1987; Battcock and Azam-Ali, 1998). There have been many reports that L. plantarum produces antimicrobial substances that inhibit food borne pathogen and spoilage microorganisms (Molin, 2001; Fooks and Gibson, 2002; Sjogren et al., 2003; Hernnadez et al., 2005). L. plantarum is recognized as a probiotic strain that strongly inhibits human intestinal pathogens such as Salmonella enteritidis and E. coli (Fooks and Gibson 2002). In addition, among the Lactobacillus strain, L. plantarum 2142 was shown to be the best producer of hydrogen peroxide and bacteriocins (Zalan et al., 2005).
Different inoculation procedures on wild forest noni fermentation: The numbers of LAB cells in the LAB and FWB sets were significantly higher than in the control set in spite of the finding that the numbers of LAB cells dropped by about 3 log cycles at day 1 of fermentation. This loss is most likely due to the LAB cells requiring time to adapt themselves to a new environment. It is also not surprising that the numbers of yeast cells were significantly less after inoculation than in the control. This may be due to a combination of having an inoculum being dominated by LAB and to the LAB producing some anti-yeast metabolites (Table 1). The higher yeast numbers in the FWB at the start probably results from their presence in the inoculum from the previous fermentation. However as the yeast would have been dominated by the LAB more rapid growth of the LAB would lead to inhibition of the yeast so that none were detected after day 14. In contrast, in the control in the presence of lower numbers of LAB the numbers of yeast increased until day 5 and to 1.6 log cfu mL-1 at the end of the fermentation (Fig. 2). This number could pass the yeast numbers set by the Thailand rules provided by the Medical Science Department, of not more than 100 cfu mL-1. However, it is well recognized that the remaining of yeast in a fermented beverage product could reduce a shelf life of the finished product due to no any treatment after finishing fermentation.
Another consequence of the inoculation procedure is that both sets of fermentations produced more acidity than that found in a control set due to the higher number of LAB cells present after inoculation, and the higher number of yeast in the control determined that the ethanol concentrations were higher in this situation. L. plantarum can also produce ethanol as it is a facultative heterofermenter and can use either the Embden-Meyerhof pathway or the phosphoketolase pathway depending on the conditions of growth (Axelsson, 2004). This explains why ethanol is produced in the LAB set in the absence of yeasts (Fig. 2) as the fermentative conditions at sometime may determine that L. plantarum uses the latter pathway and both acetic acid and ethanol are detected in the fermented beverages (Table 2). As antibacterial activity corresponded to the amount of total acidity including ethanol (Fig. 1, Table 2, 3) we had no convincing evidence that the fermentation process might result in the extraction of bacterial inhibitors from the noni fruit. However, Inoue et al. (1981) have reported that wild forest noni fruit may have some antimicrobial compounds. Now we have established that using an inoculum of a selected Lactobacillus can benefit the final product in terms of its yeast and ethanol content and ability to meet Thai health requirements.
CONCLUSION
To conclude, the results of this study show that LAB play an important role in the production of FPBs in that they produce extracellular and diffusible metabolites that inhibit potential pathogens and spoilage organisms including yeast, thereby helping to preserve and make the finished product safer. Lactic acid bacteria, such as L. plantarum W90A, originally isolated from the FWB could improve the quality of the fermented beverage produced from the fruit of wild forest noni as it completely inhibited yeast and provided more antibacterial activity. Local producers should therefore be encouraged to use standard procedures that include the use of a standard controlled inoculum for producing their FPBs.
ACKNOWLEDGMENTS
We would like to acknowledge National Science and Technology Development Agency (NSTDA), Thailand for fully support (CO-B-22-2C-18-601). We also would like to thank Dr. Brian Hodgson for many comments and suggestions.
REFERENCES
- Battcock, M. and S. Azam-Ali, 1998. Fermented Fruits and Vegetables: A Global Perspective. FAO Agricultural Services Bulletin No. 134, Food and Agriculture Organization of the United Nations, Rome, ISBN: 92-5-104226-8.
Direct Link - Chung, H.J. and A.E. Yousef, 2005. Lactobacillus curvatus produce a bacteriocin-like agent active against gram negative pathogenic bacteria. J. Food Saf., 25: 59-79.
Direct Link - Clavero, M.R. and L.R. Beuchat, 1996. Survival of Escherichia coli O157: H7 in broth and processed salami as influenced by pH, water activity and temperature and suitability of media for its recovery. Applied Environ. Microbiol., 62: 2735-2740.
Direct Link - Daeschel, M.A., R.E. Andersson and H.P. Fleming, 1987. Microbial ecology of fermenting plant materials. FEMS Microbiol. Rev., 46: 357-367.
CrossRefDirect Link - Fooks, L.J. and G.R. Gibson, 2002. In vitro investigations of the effect of probiotics and prebiotics on selected human intestinal pathogens. FEMS Microbiol. Ecol., 39: 67-75.
CrossRef - Foster, J.W., 1991. Salmonella acid shock proteins are required for the adaptive acid tolerance response. J. Bacteriol., 173: 6896-6902.
CrossRefDirect Link - Gao, Y., M.J. van Belkum and M.E. Stiles, 1999. The outer membrane of Gram-negative bacteria inhibits antibacterial activity of Brochocin-C. Applied Environ. Microbiol., 65: 4329-4333.
PubMedDirect Link - Hernández, D., E. Cardell and V. Zarate, 2005. Antimicrobial activity of lactic acid bacteria isolated from Tenerife cheese: Initial characterization of plantaricin TF711, a bacteriocin-like substance produced by Lactobacillus plantarum TF711. J. Appl. Microbiol., 99: 77-84.
CrossRefPubMedDirect Link - Inoue, K., H. Nayeshiro, H. Inouye and M. Zenk, 1981. Anthraquinones in cell suspension cultures of Morinda citrifolia. Phytochemistry, 20: 1691-1700.
CrossRefDirect Link - Ivanova, I., P. Kabadjova, A. Pantev, S. Danova and X. Dousset, 2000. Detection, purification and partial characterization of novel bacteriocin substance produced by Lactococcus lactis subsp. lactis B14 isolated from Boza-Bulgarian traditional cereal beverage. Biocatal. Fundam. Appl., 41: 47-53.
Direct Link - Kang, D.H. and D.Y. Fung, 1999. Effect of diacetyl on controlling Escherichia coli O157: H7 and Salmonella Typhimurium in the presence of starter culture in a laboratory medium and during meat fermentation. J. Food Protect., 62: 975-979.
CrossRefDirect Link - Kantachote, D., W. Charernjiratrakul and N. Aussawariangpipop, 2005. Characteristics of fermented plant beverages in Southern Thailand. Songklanakarin J. Sci. Technol., 27: 601-615.
Direct Link - Kantachote, D., M. Ongsakol, W. Charernjiratrakul, C. Chaiyasut and N. Poosaran, 2005. Fermented plant beverages and their application. Proceedings of the 1st International Conference on Natural Products for Health and Beauty from Local Wisdom to Global Marketplace, Oct. 17-21, Maha Sarakham, Thailand.
- Kantachote, D. and W. Charernjiratrakul, 2007. Effects of initial air removal methods on microorganisms and characteristics of fermented plant beverages. Pak. J. Biol. Sci., 11: 173-180.
CrossRefPubMedDirect Link - Lindgren, S.E. and W.J. Dobrogosz, 1990. Antagonistic activities of lactic acid bacteria in food and feed fermentations. FEMS Microbiol. Rev., 7: 149-163.
PubMedDirect Link - Molin, G., 2001. Probiotics in foods not containing milk or milk constituents, with special reference to Lactobacillus plantarum 299v. Am. J. Clin. Nutr., 73: 380s-385s.
CrossRefDirect Link - Osmanagagaoglu, O., F. Kiran and N. Gul, 2005. Effect of pediocin DT10 on Leuconostoc mesenteroides OZ-N3 cells. J. Food Saf., 25: 303-317.
CrossRefDirect Link - Prachyakij, P., J. Schnurer, W. Charernjiratrakul and K. Kantachote, 2007. Selection and identification of lactic acid bacteria that inhibit yeast contaminants isolated from fermented plant beverages. Songklanakarin J. Sci. Technol., 29: 211-218.
Direct Link - Schillinger, U. and F.K. Lucke, 1989. Antibacterial activity of Lactobacillus sake isolated from meat. Applied Environ. Microbiol., 55: 1901-1906.
Direct Link - Simango, C. and G. Rukure, 1992. Survival of bacterial enteric pathogens in traditional fermented foods. J. Applied Bacteriol., 73: 37-40.
CrossRefDirect Link - Sjogren, J., J. Magnusson, A. Broberg, J. Schnurer and L. Kenne, 2003. Antifungal 3-hydroxy fatty acids from Lactobacillus plantarum milab 14. Applied Environ. Microbiol., 69: 7554-7557.
CrossRefDirect Link - Spelhaug, S.R. and S.K. Halander, 1989. Inhibition of foodborne bacterial pathogens by bacteriocins from Lactococcus lactis and Pediococcus pentosaceous. J. Food Protect., 52: 856-862.
CrossRefDirect Link - Yang, M. and Y. Choong, 2001. A rapid gas chromatographic method for direct determination of short-chain (C1-C12) volatile organic acids in foods. Food Chem., 75: 101-108.
Direct Link - Yap, P.S. and S.E. Gilliland, 2000. Comparison of newly isolated strains of Lactobacillus delbrueckii subsp. lactis for hydrogen peroxide production at 5�°C. J. Dairy Sci., 83: 628-632.
Direct Link - Zalan, Z., E. Nemeth, A. Barath and A. Halasz, 2005. Influence of growth medium on hydrogen peroxide and bacteriocin production of Lactobacillusv strains. Food Technol. Biotechnol., 43: 219-225.
Direct Link