
Vahid Najati
Faculty of Sciences, Urmia University, Urmia, Iran
Minoo Ilkhanipour
Faculty of Sciences, Urmia University, Urmia, Iran
Shahpar Salehi
Faculty of Sciences, Urmia University, Urmia, Iran
Goudarz Sadeghi-Hashjin
Faculty of Veterinary Medicine, University of Tehran, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 2 | Page No.: 250-254







ABSTRACT
The aim of this study was to investigate the role of Nitric Oxide (NO) in the atresia of ovarian follicles in an animal model. Twenty adult, female rats (90 days old with body weights of 210 ± 10 g in the beginning of the experiments) were divided into 4 groups of 5 each. They were treated twice daily from the subcutaneous route for 21 successive days with either of the following chemicals: nitroglycerine, L-arginine, L-NAME, or saline. On day 22, all animals were sacrificed. Ovaries were dissected out free of connected tissue and were fixed in formaline 10%. Later, paraffine blocks were prepared and serial sections were made by means of H and E routine staining method. Intact and atretic follicles were counted separately. In addition, damages were analyzed qualitatively from the points of view of appearance and morphologic changes. In the evaluation of ovarian follicular structures, different types of healthy as well as atretic follicles were observed. In most of atretic follicles, the oocytes were abnormally elongated and increnation of their outlines were obvious. There were numerous macrophages around and inside of the atretic follicles. Our investigation regarding the distribution of atretic follicles in the ovaries of test groups revealed that atretic follicles in the L-NAME treated group were increased in comparison to the control group. Conversely, however, in the arginine-treated group, the atretic follicles were reduced compared to the control animals. Treatment with nitroglycerine of the rats decreased the number of atretic follicles significantly (p<0.05) in comparison to the control group. In conclusion, enhanced NO, either from endogenous or exogenous origins, prevents atresia phenomenon, while inhibition of NO exerts an opposite effect.
PDF Abstract XML References Citation
How to cite this article
Vahid Najati, Minoo Ilkhanipour, Shahpar Salehi and Goudarz Sadeghi-Hashjin, 2008. Role of Nitric Oxide on the Generation of Atretic Follicles in the Rat Ovaries. Pakistan Journal of Biological Sciences, 11: 250-254.
DOI: 10.3923/pjbs.2008.250.254
URL: https://scialert.net/abstract/?doi=pjbs.2008.250.254
DOI: 10.3923/pjbs.2008.250.254
URL: https://scialert.net/abstract/?doi=pjbs.2008.250.254
INTRODUCTION
Nitric Oxide (NO) is a chemical mediator present in different tissues as in human reproductive system (Calka, 2006). It is involved in the regulation of many functions in the reproductive system, including parturition, pregnancy, implantation, ovular activities, steroidogenesis, folliculogenesis and the regulation of LHRH (Jain et al., 1993). NO was reported to increase the levels of LH significantly with no effect on the production of prolactin (Russell et al., 2005). In addition, decreased levels of NO enhanced the contraction of smooth muscle in the fallopian tubes, hence, increasing egg transport in the ovum (Ekerhord et al., 1999). NO increases endometrial secretions during the oogenesis phase in female animals (Morlin and Hammarstrom, 2005), increases blood circulation and plays a role in follicle development, oocyte maturation and steroidogenesis (Philip et al., 2001).
In this study, we investigated the role of this putative mediator on follicular atresia in the rat. For this purpose, animals were treated with either of the following substances: saline (control), nitroglycerine (NTG, an NO donor), L-arginine (LARG, precursor of NO), or N[g]-nitro-L-arginine methyl ester (L-NAME, an inhibitor of NO synthase [NOS]).
MATERIALS AND METHODS
This study was performed in the year 2006 in Urmia University Faculty of Sciences, Urmia, Iran.
Animals: Twenty adult, female rats were used in this study. They were 90 days old, with body weights of 210±10 g in the beginning of the experiments. They were fed with commercial chow and tap water ad lib and kept at room temperature with 12 h artificial light in 24 h.
Experimental protocol: Animals were divided into 4 groups of 5 each. They were treated twice daily from the subcutaneous route for 21 successive days with either of the following chemicals: Group A with nitroglycerine (an NO donor), group B with L-arginine (the precursor of NO), group C with L-NAME (an inhibitor of NO synthase) and group D with saline solution (control). On day 22, all animals were sacrificed. Ovaries were dissected out free of connected tissue and were fixed in formaline 10%. Later, paraffine blocks were prepared and serial sections were made by means of H and E routine staining method. Intact and atretic follicles were counted separately. In addition, damages were analyzed qualitatively from the points of view of appearance and morphologic changes.
Chemicals: L-NAME, LARG and NTG were products of Sigma (St. Luis, USA), Merck (Darmstadt, Germany) and Daroupakhsh Pharmaceutical Company (Tehran, Iran), respectively.
Statistics: Differences in the number of atretic follicles were first analyzed by one-way ANOVA and thereafter by Bonferroni`s t-test. A p-value smaller than 0.05 was considered as the level of significance.
RESULTS
In the evaluation of ovarian follicular structures, different types of healthy as well as atretic follicles were observed. The number of atretic follicles were enhanced slightly after L-NAME treatment while L-arginine and nitroglycerine had an opposite effect; the effect of the latter reached the level of significance (p<0.05, Fig. 1). In most of atretic follicles, the oocytes were abnormally elongated and increnation of their outlines were obvious. Follicles with 2-3 granulosa cells were exhibiting dispersed antral spaces between them (precautions antral formation) as a sign of follicular atresia. In some primary and secondary follicles, signs of luteinization were seen in granulosa cells, which is also a potential sign of follicular atresia. In atretic follicles pyknotic index in granulose cells were high, further revealing follicular atresia. There were numerous macrophages around and inside of the atretic follicles; by the progression of follicular atresia, their population increased accordingly. The structure of Zona Pleucida (ZP) was affected in atretic follicles and signs of breakdown perforation and fragmentation were seen in ZP structure of atretic follicles. Present investigation regarding the distribution of atretic follicles in the ovaries of test groups revealed that atretic follicles in the L-NAME treated group were increased in comparison to the control group. Conversely, however, in the arginine-treated group, the atretic follicles were reduced compared to the control animals. Treatment with nitroglycerine of the rats decreased the number of atretic follicles significantly (p<0.05) in comparison to the control group (Fig. 1). In the atretic follicles, the following changes were observed: cleavage-like divisions in oocyte nucleus (Fig. 2) and malformed oocyte with shrinkage and luteinized granulosa follicle (Fig. 3).
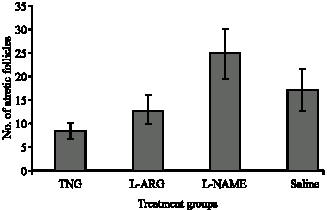 | |
Fig. 1: | Comparison of the avenge numbers of atretic follicles between control and treated experimental groups. As seen in the figure, increased levels of NO by L-Arg and TNG (p<0.05) decreased the number of atretic follicles, while inhibition by L-NAME of NO production had the opposite effect |
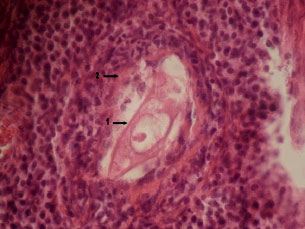 | |
Fig. 2: | Oocyte nucleus shows cleavage-like divisions (1) and a macrophage is seen inside the entrum |
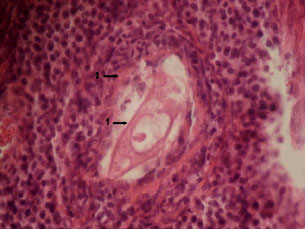 | |
Fig. 3: | Arrow number 1 shows a malformed oocyte with shrinkage and number 2 demonstrates granulosa follicle that has been luteinized |
DISCUSSION
Although research activities have long been done over follicular atresia, the underlying mechanisms are not fully understood yet. Whilst some believe that early signs are observed in the oocyte, others claim that it is initiated from granulosa cells and then spread to other parts (Asami, 1920; Sturgis, 1949; Ingram, 1962; Pincus and Enzman, 1975; Daud et al., 1988). Follicular atresia is a process which may occur during any stage of the follicular growth (Byskov and Rasmussen, 1974). Differential diagnosis between normal and atretic follicles is a difficult task as all follicles do not follow a similar process during the phenomenon (Byskov, 1979; Grimes et al., 1987).
Of numerous number of the follicles in an ovary, only a limited number succeed in growth, maturity and, finally, ovulation. The rest become atretic. Atresia may occur in different forms, including picnosis of granulosa cells (Hirshfield, 1989), segmentation of basal layer (Bagavandoss et al., 1983), floating the granulosa cells in the antral fluid (Hay et al., 1976), shrinkage, segmentation and disappearance of the nucleus and nucleolus (Ojeda et al., 1992; Hirshfield and Midyley, 1978; Peters and Ball, 1987).
Various forms of atresia were observed in the present study. The interesting finding, however, was that many atretic follicles possessed a few characteristics of atresia so that, especially in larger follicles, more than 6 important signs of atresia phenomenon were observed.
In cytologic criteria, picnosis and chromatinization of granulosa cells, disrupture between them and existence of floating granulosa cells in the antrum with picnotic nuclei are evident signs of follicular atresia (Byskov and Rasmussen, 1974).
During recent years, studies have shown that macrophages have a great impact on the regulation of the ovarian functions (Wu et al., 2004). They reported that macrophages and their secretory products can lead to disturbances in the ovaries, including polycystic ovary syndrome, endometriosis and early failure of ovaries. In addition, the role of these cells in the destruction of cellular remnants in various tissues of the body has already been shown. Therefore, it can be concluded that in the ovarian tissue, whenever atresia or cell death occurs, macrophages are also present.
In this study, too, the presence of macrophages was shown in different areas of the ovarian tissue. No macrophages were seen around healthy follicles but they had an active presence around atretic ones. It was possible to locate many macrophages in the antrum of antral follicles and many of them were seen in the connective tissue surrounding non-healthy (atretic and cystic) follicles. Following ovulation, the cellular complex of the follicles undergoes alterations that finally lead to the formation of the corpus luteum (Byskov, 1979). The granulosa and internal tech participate in the formation of corpus luteum. The granulosa cells are larger and paler while the thecal cells are smaller and denser than the others (Dellman and Eurell, 1998). Based on what was said, it can be concluded that any luteinization of the follicular wall that occurs before ovulation is a non-physiologic process and can be considered as atretic processes.
Some other important signs of atresia may be manifested in the oocyte of which the mitotic division and cleavage-like changes and segmentation of oocyte are common (Talukar et al., 1991). Similar changes were observed in this study as well. Mitotic and cleavage-like divisions were demonstrated in the follicles larger than 200 μm. We concluded that these abnormalities are seen solely in tertiary and graffian follicles, whereas small follicles at early stages of growth were free of these disturbances.
Shrinkage and malformation of the oocyte are other prominent signs of atresia (Hirshfield, 1989). In normal follicles, oocytes are spherical with a round shape seen in their cross sections. The nucleus is located in the centre and is seen clearly together with the nucleolus and the oocyte has a completely smooth wall. However, we observed a number of oocytes with serrated and rough surroundings. Oocyte was crumpled and shrunken form. Surroundings of the oocyte had a shape of serrated, folded and dented appearance. In most cases, this cell was detached from the granulosa cells and from its cumulous mass in the antral follicles and was floating in the follicular fluid. This abnormality was observed in all stages of follicular growth, including antral and pre-antral stages.
It should be stated that any change in the structure of ZP can affect the health of oocyte, but it should be also mentioned that degenerative changes in the ZP take place following degenerative changes in the oocytes and granulosa cells (Hasanzadeh and Sadrkhanloo, 2002). The present research work demonstrated several cases of atresia, related to ZP. This abnormality was seen in the follicles of more progressive stage. From the disturbances seen in the ZP, the followings can be numbered: deletion of some parts of the membrane, perforation, laminating, hyalinization, breakage, incarnation and folding in ZP.
Studies performed on the role of NO in the rabbit shows that this substance has effects on ovulation, oocyte maturation and production of sex steroids. By relaxing the vascular smooth muscle, NO increases blood flow in the ovary, easing the rupture of the follicle and ovulation. In addition, endogenous NO and NO-related substances inhibit steroid production by the luteal and granulosa cells (Yamauchi et al., 1997). In vitro studies suggest a clear role for NO in the persistence of the ovum as well as on ovulation. The effects of NOS inhibitors in the body, however, are not identical with those seen under in vitro conditions; it is possible that these effects in the body are mediated by extra-follicular factors as well (Mitchel et al., 2004). After ovulation, homogenous endothelial NOS can be seen in corpus luteum. During investigations performed on mouse ovary, it was observed that eNOS-derived NO is the mediator of oocyte miotic maturation. Besides, NO is involved in the PSRAFT (regression) of the luteal function via inhibition of steroidogenesis. NO may induce rupture of follicles in the rabbit during the ovulation process and this is probably accomplished via prostaglandin production (Tsafriri et al., 1996).
Based on what seen in the present findings, L-NAME and L-arginine, respectively, have dramatically increased and decreased the number the atretic follicles. Other findings have revealed that NO is significantly implicated in apoptosis. NO may decrease apoptosis by nitrosylating cysteine and hence, inhibition of caspase-3 (Lothar and Cherer, 1999). Studies have demonstrated that endogenous NO, produced by granulosa cells of immature follicles, may decline apoptosis and, therefore, prevent atresia development in follicles. Indeed, elevated levels of NO have been shown to decrease apoptosis in the granulose cells in pigs and calves (Tamanini et al., 2003). This could be one of the underlying mechanisms to decline atresia of the follicles in the animal model used in this study.
REFERENCES
- Asami, G., 1920. Observations on the follicular atresia in the rabbit ovary. Anat. Rec., 18: 323-343.
CrossRef - Bagavandoss, P., A.R.J. Midgley and M. Wicha, 1983. Developmental changes in the ovarian follicular basal lamina detected by immunofluorescence and electron microscopy. J. Histochem. Cytochem., 31: 633-640.
Direct Link - Calka, J., 2006. The role of nitric oxide in the hypothalamic control of LHRH and oxytocine release, sexual behavior and aging of the LHRH and oxytocine neurons. Folia Histochem. Cytobiol., 44: 3-12.
Direct Link - Daud, A.L., F.M. Bumpus and A. Husain, 1988. Evidence for selective expression of angiotensin 2 receptors on atretic follicle in the rat ovary and auto radiographic study. Endocrinology, 122: 2727-2734.
CrossRef - Ekerhord, E., M. Brannstrom, B. Weijdegard and A. Norstrom, 1999. Localization of nitric oxide synthase and effects of nitric oxide donors on the human fallopian tube. Mol. Hum. Reprod., 5: 1040-1041.
Direct Link - Grimes, R.W., P. Matton and J.J. Irland, 1987. A comparison of histological and non-histological indices of atresia and follicular function. Biol. Reprod., 37: 82-88.
CrossRef - Hay, M.F., P.G. Cran and R.M. Moor, 1976. Structural changes occurring during atresia in sheep ovarian follicles. Cell Tissue Res., 169: 515-529.
CrossRef - Hirshfield, A.N. and A.R. Midyley, 1978. Morphologic analysis of follicular development in the rat. Biol. Reprod., 190: 597-605.
CrossRef - Hirshfield, A.N., 1989. Rescue of atresic follicles in vitro and in vivo. Biol. Reprod., 40: 181-190.
PubMed - Jain, B., I. Rubinstein, R.A. Robbins, K.L. Leise and J.H. Sission, 1993. Modulation of airway epithelial cell ciliary beat frequency by nitric oxide. Biochem. Biophys. Res. Commun., 191: 83-88.
PubMed - Lothar, R. and B.F. Cherer, 1999. Nitric oxide inhibits caspase-3 by s-nitrosation in vivo. J. Biol. Chem., 274: 6823-6826.
PubMed - Mitchel, L.M., C.R. Kennedy and G.M. Hartshorne, 2004. Pharmacological manipulation of nitric oxide levels in mouse follicle cultures demonstrates key role of extra follicular control of ovulation. Hum. Reprod., 19: 1705-1712.
Direct Link - Morlin, B. and M. Hammarstrom, 2005. Nitric oxide increases endocervical secretion at the ovulatory phase in the female. Acta obstetricia Gynecol. Scandinavica, 84: 883-886.
Direct Link - Philip, P.M.W., R. Pathansali, R. Iddenden and F. Bath, 2001. The effect of transdermal glyceryl trinitrate, a nitric oxide denor, on blood pressure and platelet function in acute stroke. Cerebrovasc Dis., 11: 265-272.
Direct Link - Pincus, G. and E.V. Enzman, 1975. The growth, maturation and atresia of ovarian eggs in the rabbit. Morphology, 61: 351-383.
CrossRef - Russell, J.M., E. Murphree, J. Janik and P. Callahan, 2005. Effect of steroids and nitric oxide on pituitary hormone release in ovariectomized, peripubertal rats. Reproduction, 129: 497-504.
Direct Link - Sturgis, H.S., 1949. Rate and Significance of atresia in the ovarian follicle of the rhesus monkey. Embryol. Corneg. Inst., 33: 67-70.
PubMed - Tsafriri, A., S.Y. Chun, A.J.W. Hsueh and M. Conti, 1996. Oocyte maturation involves compartmentalization and opposing changes of cAmp levels in follicular somatic and germ cells: Studies using selective phosphodiesterase inhibitors. Dev. Biol., 178: 393-402.
CrossRef - Wu, R., K.H. Vander Hock, N.K. Rayan, R.J. Norman and R.L. Robker, 2004. Macrophage contribution to ovarian function. Hum. Reprod. Update, 10: 113-119.
Direct Link - Yamauchi, J., T. Miyuzaki, S.H. Iwasaki, H. Kishi, M. Kuroshima, C.H. Tei and Y. Yoshimura, 1997. Effect of nitric oxide on ovulation and ovarian steroidogenesis and prostaglandin production in the Rabbit. Endocrinology, 138: 3630-3637.
CrossRef