
K.J. Khorshidi
Department of Animal Science, Islamic Azad University, Ghaemshahr Branch, Iran
A. Karimnia
Department of Animal Science, Islamic Azad University, Ghaemshahr Branch, Iran
S. Gharaveisi
Department of Animal Science, Islamic Azad University, Ghaemshahr Branch, Iran
H. Kioumarsi
School of Biological Sciences, Universiti Sains Malaysia, Penang, Malaysia
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 20 | Page No.: 2395-2400







ABSTRACT
This research was conducted in order to investigate the effects of supplemental fat and monensin on the level of Dry Matter Intake (DMI), Average Daily Gain (ADG), final weight, Feed Conversion Ratio (FCR), blood glucose level, Blood Urea Nitrogen (BUN), calcium, phosphor, triglyceride, cholesterol and crude protein of the Zel lambs; as well as commercial effects of monensin and supplemental fat on the lambs fattening. Most of the time, ordinary food does not suffice the animals needs. Thus, using high-energy materials like fats are important in programming the diets. Moreover, adding some materials to the animal diets could increase the animal efficiency and the products quality. Hormones, antibiotics, ionophores are amongst the most important additives affecting income in the lamb fattening industry. Monensin is the first ionophore component proven to be effective in increasing efficiency of ruminant animals. Twenty-four male Zel lambs were used with an age average of 3-4 months and an initial live weight of 21.94 ± 0.642 kg. A period of eleven days was considered as the adaptation time and the experiment was carried on during three 21-day phases (totally 63 days). The variables for this study were supplemental fat on two levels of 0 and 4% and monensin on tree levels of 0, 20 and 40 mg kg-1 on adrymatterbasis. There were four repetitions for each treatment. The results of this research reveal that different levels of fat and monensin do not have significant effect on DMI, ADG, FCR and level of glucose. The food treatment had significant effect on final weight, calcium level, BUN, phosphor, triglyceride, cholesterol and CP (p<0.05). Also, they affect commercial productivity.
PDF Abstract XML References Citation
How to cite this article
K.J. Khorshidi, A. Karimnia, S. Gharaveisi and H. Kioumarsi, 2008. The Effect of Monensin and Supplemental Fat on Growth Performance, Blood Metabolites and Commercial Productivity of Zel Lamb. Pakistan Journal of Biological Sciences, 11: 2395-2400.
DOI: 10.3923/pjbs.2008.2395.2400
URL: https://scialert.net/abstract/?doi=pjbs.2008.2395.2400
DOI: 10.3923/pjbs.2008.2395.2400
URL: https://scialert.net/abstract/?doi=pjbs.2008.2395.2400
INTRODUCTION
Ionophores are an associate polyether antibiotic group and are used as growth promoters in ruminants and pigs. They are used as a coccidiostat in poultry and ruminants (McKellar and Lawrence, 1996; Salles et al., 2008). Antibiotic ionophores have been widely used to improve feed efficiency and to regulate rumen functions in animals. The ionophore decreases methanogenesis (Thornton and Owens, 1981; Crutzen et al., 1986; Rumpler et al., 1986; Mbanzamihigo et al., 1996; Guan et al., 2006) and increases the molar proportion of propionate while acetate and often butyrate proportions are decreased (Horn et al., 1981; Broderick, 2004). In addition, they act by increasing Non-Ammonia Nitrogen (NAN) flows in the duodenum of treated animals (Schelling, 1984; Rogers et al., 1997). Monensin is an ionophore produced by a Streptomyces cinnamonensis sub-type and improves animal feed efficiency. On the other hand, the most important reason for using supplemental fat in ratio is the increase of dietary energy level without the increase of starch in using diet. Therefore, it can be assumed that we can obtain increase in dietary energy level without any change in Dry Matter Intake (DMI) after adding fat. Soodeen-Karamath and Youssef (1999) assessed the effect of monensin, avoparcin and grass supplementation on utilization of urea-treated rice straw by sheep and goats and concluded that diets based on urea-treated rice straw supplemented with grass and/or monensin were utilized more efficiently by sheep than goats. Garcia et al. (2000) assessed the effect of a yeast culture (Saccharomyces cerevisiae) and monensin on ruminal fermentation and digestion in sheep and showed that although supplementing sheep with a combination of S. cerevisiae and monensin had no effect on digestibility or ruminal fermentation, monensin improved the fermentation pattern. According to Clary et al. (1993), supplemental fat and ionophores can be bound and as a result, the amount of inserting fat to intestine will be increased. Wholly, using supplemental fat and ionophores can be as a strong method to improve ruminant`s nutrition and there have been many other researches by the scientist on the different species in this regard. Nonetheless, the data does not seem to be adequate yet. This study was carried out to evaluate the effect of monensin and supplemental Fat on growth performance, blood metabolites and commercial productivity of Zel lambs.
MATERIALS AND METHODS
Lambs and experimental conditions: This study was conducted in 2007 as a portion of Animal Science Research Program in Agricultural and Animal Science Research Center of Azad University, Ghaemshahr Branch in north of Iran. Twenty-four male Zel lambs were used with an age average of 6 months and an initial live weight of 21.94 ± 0.642 kg. The animals were allowed 50 days period to adjust to the new feeding and housing conditions prior to the start of the experiment. The animals were housed in experimental pens and fed two times a day with Total Mix Ratio (TMR) for a period of 63 days including 3 periods and each period lasted 21 days.
Experimental diets: Chemical analysis of the feeds was performed according to AOAC (1990). Subsequently, with reference to the average age and weight of the lambs and based on the standards set by the NRC, six food rations were prepared with two levels of Fat (0 and 4%) and one of three levels of monensin (0, 20 and 40%). Food was offered in the morning and evening and the food refused from the previous day was removed before the new meal was given on the following morning.
Blood metabolites: Blood samples were collected from the jugular veins. All blood samples were stored in ice in a sterile vacutainer that contained EDTA as an anti-coagulant. In the lab, the blood samples were centrifuged at 5,000 x G for 6 min. Subsequently, using commercial kits, the amounts of glucose, urea, calcium, phosphor, triglyceride and cholesterol based on mg/100 mL and Crude Protein (CP) based on g 100 mL-1 in the samples were determined.
Statistical analysis: A completely randomized design of 6 diets in a 2x3 factorial design was replicated four times. The model was:
Xijk = M+Bj+Bk+Bjk+Eijk |
Where:
| Xijk | = | Observations |
| M | = | Treatment average |
| Bj | = | Effect of fat level |
| Bk | = | Effect of monensin level |
| Bjk | = | Effect of the interaction between fat and monensin levels |
| Eijk | = | Experimental error |
Data were statistically analyzed using the General Linear Model procedures of SAS (1996) with the Duncan test.
RESULTS
Growth performance: The fat and monensin levels were found to have a significant effect on final weight (p<0.05). The ration containing 4% fat and 20 mg kg-1 Dry Matter (DM) monensin resulted in the most favorable final weight. Monensin and fat did not significantly affect Dry Matter Intake (DMI), Average Daily Gain (ADG), as well as Feed Conversion Ratio (FCR) (Table 1).
Blood metabolites: Monensin and fat did not significantly affect blood glucose. The fat and monensin levels were found to have a significant effect on BUN (p<0.05) that the lowest and the highest amount was 10.33 and 19.58 mg/100 mL for lambs fed with ratios two and six, respectively (Table 2). Furthermore, the fat and monensin levels were found to have a significant effect on calcium (p<0.05) that the lowest and the highest amount was 9.60 and 10.61 mg/100 mL for lambs fed with ratios one and six, respectively. Moreover, the fat and monensin levels were found to have a significant effect on phosphor (p<0.05) that the lowest and the highest amount was 6.70 and 9.69 mg/100 mL for lambs fed with ratios six and three, respectively. In addition, the fat and monensin levels were found to have a significant effect on triglyceride (p<0.05) that the lowest and the highest amount was 15.25 and 29.25 mg/100 mL for lambs fed with ratios six and three, respectively. Also, the fat and monensin levels were found to have a significant effect on cholesterol (p<0.05) that the lowest and the highest amount was 28.91 and 51.16 mg/100 mL for lambs fed with ratios two and six, respectively. Besides, the fat and monensin levels were found to have a significant effect on CP (p<0.05) that the lowest and the highest amount was 6.24 and 7.67 g/100 mL for lambs fed with ratios three and five, respectively.
| Table 1: | Effects monensin and supplemental fat on lamb performance and efficiency |
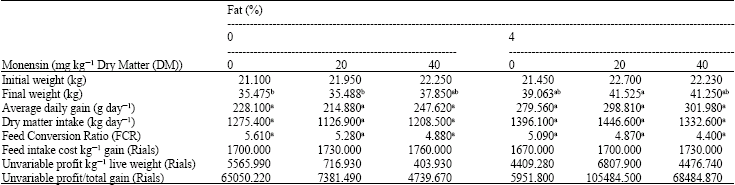 | |
| In each line, means with different letter(s) are significantly different (p<0.05) | |
| Table 2: | Effects monensin and supplemental fat on lamb blood metabolites |
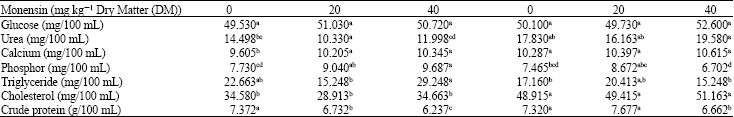 | |
| In each line, means with different letter(s) are significantly different (p<0.05) | |
Commercial productivity: The ration containing 40 mg kg-1 Dry Matter (DM) monensin and 4% fat resulted in the most favorable feed intake cost/kg gain, un-variable profit/kg live weight and un-variable profit/total gain (Table 1).
DISCUSSION
Growth performance: The addition of monensin and supplemental fat had significant effect on final weight but had no significant effect on Dry Matter Intake (DMI), Average Daily Gain (ADG), as well as Feed Conversion Ratio (FCR). Huston et al. (1990), Stock et al. (1995) and Rowghani et al. (2006) using monensin caused decrease in DMI but here the result can be justified with using fat beside monensin in order not to have any negative effects on DMI. Monensin and supplemental fat improved feed conversion ratio and increased daily weight gain, but the effects were nonsignificant. Also, it had significant effect on final weight. Ruminal fermentation has been manipulated with different feed additives, including ionophores, to improve animal production (Wallace, 1994; Garcia et al., 2000). The main described effects of monensin on rumen fermentation have been explained partially by the reduction of Gram-positive bacteria in the rumen (Russell, 1987; Devant et al., 2007) and are the increase of propionate production, the decrease in methane production (McGinn et al., 2004; Devant et al., 2007) and the increase of final weight as a result. Spears and Harvey (1984) suggested that using ionophores can create component with some cations like sodium and potassium in rumen and they will change rumen microbial activity pH and as a result, they will change absorption, metabolite process and nutrition productivity in animals. Moreover, in parallel with these changes, especially nutrition productivity, Feed Conversion Ratio (FCR) will be improved.
Blood metabolite: The effect of monensin and supplemental fat on blood glucose level was nonsignificant. Spears and Harvey (1984) assessed serum characteristics of steers fed lasalocid on pasture and observed that an increase (p<0.05) in plasma was observed in steers fed 300 mg lasalocid/day as compared with the 200 mg level and described that if the increased plasma glucose concentrations were due to an increased availability of propionate for gluconeogenesis, it is difficult to rationalize why glucose was increased only at the high level of lasalocid while propionate was similar for steers fed 200 or 300 mg lasalocid/day. Thonney et al. (1981) also did not witness any regular effects on blood glucose level in steers fed by lasalocid. Arieli et al. (2001) showed that the glucogenic effect of monensin might be only revealed in cows that are in a negative energy balance. The findings of this experiment reveal that monensin alone decreases the level of blood urea and the justification is that monensin reduced the degradation of dietary protein in the rumen, so it could be expected that blood urea nitrogen would decline (Hayes et al., 1996). On the other hand, the supplemental fat increases, as well as the simultaneous increase in monensin and supplemental fat raise the Blood Urea Nitrogen (BUN). Increasing the energy level allows the production of more fermentable ME for paunch microorganisms resulting in a rise in the synthesis of microbial protein and in the amount of protein available to the animal (Webster, 1994; Early et al., 2001; Kioumarsi et al., 2008a) and consequently, it can increase the amount of Blood Urea Nitrogen (BUN). Adding fat and monensin to the diet has significant effect on the level of calcium and phosphorous in increasing them. Monensin modifies the flux of ions across intestinal epithelial cells and increase uptake and availability of dietary minerals (Beckett et al., 1998; Kirk et al., 1985a, b).
Supplemental fat and monensin have significant effect on the blood triglyceride level. Yang et al. (2003) stated that monensin has significant effect on the blood triglyceride level of the goat fed by monensin. By adding supplemental fat, more fatty acids enter the intestine and thus, absorbed. Triglycerides are needed to transfer the absorbed fatty acids in the blood. As a result, the level of blood triglycerides increases. By adding fat to the diet number four, the level of cholesterol significantly increase. At the same time, by adding fat and monensin simultaneously (diets five and six) the level of cholesterol increases significantly compared to that of the control diet. Studying the effects of fatty acids produced by saturated acids, the researchers have observed that adding these fats to the diet would increase the blood cholesterol level. They also found that the change in the blood cholesterol level is due to changes occurring in the liver. When the saturated fatty acids enter the liver, cholesterol goes into the regulatory pool and gets out of the ester pool. This decreases the activity level of LDL receptor in the liver, that is to say the connecting factor to the cholesterol and the one that transfers it decrease and thus, the produced cholesterol receptor increases (Drackley et al., 1992, 1998; Mohamed et al., 1998; Staples et al., 1997).
Commercial productivity: This experiment reveals that increase in monensin and supplemental fat to diet would improve the economical productivity. A number of factors affect productivity in sheep marketing, of which the dietary energy and protein levels and their interaction are probably the most important (Muwalla et al., 1998; Bellof and Pallauf, 2004; Kioumarsi et al., 2008b). Supplemental fat will increase dietary energy level and increasing the energy level allows the production of more fermentable ME for paunch microorganisms resulting in a rise in the synthesis of microbial protein and in the amount of protein available to the animal (Early et al., 2001; Kioumarsi et al., 2008a). On the other hand, monensin is an ionophore that improves animal feed efficiency (Salles et al., 2008).
CONCLUSION
In this research, it was observed that adding monensin and supplemental fat improve feed conversion ratio, increases daily weight gain and consequently increase the final weight. Moreover, it was observed that adding monensin alone decreases the dry matter intake, though it will increase it when accompanied by fat. It also has significance on blood metabolite. On the whole, although these effects on the feed conversion ratio and daily weight gain are nonsignificant, they surely tap on commercial productivity.
REFERENCES
- AOAC, 1990. Official Methods of Analysis. 15th Edn., Association of Official Analytical Chemists, Washington, DC., USA., pp: 200-210.
Direct Link - Arieli, A., J.E. Vallimont, Y. Aharoni and G.A. Varga, 2001. Monensin and growth hormone effects on glucose metabolism in the prepartum cow. J. Dairy Sci., 84: 2770-2776.
Direct Link - Beckett, S., I. Lean, R. Dyson R, W. Tranter and L. Wade, 1998. Effects of monensin on the reproduction, health and milk production of dairy cows. J. Dairy. Sci., 81: 1563-1573.
Direct Link - Bellof, G. and J. Pallauf, 2004. Deposition of protein, fat and energy in lambs of the breed German Merino Landsheep. Anim. Sci., 78: 369-378.
Direct Link - Broderick, G.A., 2004. Effect of low level monensin supplementation on the production of dairy cows fed alfalfa silage. J. Dairy Sci., 87: 359-368.
Direct Link - Guan, H., K.M. Wittenberg, K.H. Ominski and D.O. Krause, 2006. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci., 84: 1896-1906.
Direct Link - Clary, E.M., R.T. Brandt Jr., D.L. Harmon and T.G. Nagaraja, 1993. Supplemental fat and ionophores in finishing diets: Feedlot performance and ruminal digesta kinetics in steers. J. Anim. Sci., 71: 3115-3123.
Direct Link - Crutzen, P.J., I. Aselmann and W. Seiler, 1986. Methane production by domestic animals, wild ruminants, other herbivorous fauna and humans. Tessus, 38B: 271-284.
Direct Link - Drackley, J.K., D.W. LaCount, J.P. Elliott, T.H. Klusmeyer and T.R. Overton et al., 1998. Supplemental fat and nicotinic acid for Holstein cows during an entire lactation. J. Dairy Sci., 81: 201-214.
Direct Link - Devant, M., A. Anglada and A. Bach, 2007. Effects of plant extract supplementation on rumen fermentation and metabolism in young Holstein bulls consuming high levels of concentrate. Anim. Feed. Sci. Technol., 137: 46-57.
CrossRefDirect Link - Early, R.J., O. Mahgoub and C.D. Lu, 2001. Energy and protein utilization for maintenance and growth in Omani ram lambs in hot climates. II. Composition of tissue growth and nitrogen metabolism. J. Agric. Sci., 136: 461-470.
CrossRefDirect Link - Garcia, C.C.G., M.G.D. Mendoza, M.S. Gonzales, P.M. Cobos, C.M.E. Ortega and L.R. Ramirez, 2000. Effect of a yeast culture (Saccharomyces cerevisiae) and monensin on ruminal fermentation and digestion in sheep. Anim. Feed. Sci. Technol., 83: 165-170.
Direct Link - Hayes, D.P., D.U. Pfeiffer and N.B. Williamson, 1996. Effect of intraruminal monensin capsules on reproductive performance and milk production of dairy cows fed pasture. J. Dairy Sci., 79: 1000-1008.
Direct Link - Horn, G.W., T.L. Mader, S.L. Armbruster and R.R. Frahm, 1981. Effect of monensin on ruminal fermentation, forage intake and weight gains of wheat pasture stocker cattle. J. Anim. Sci., 52: 447-454.
Direct Link - Huston, J.E., B.S. Engdhal and M.C. Calhoun, 1990. Effect of supplemental feed with or without ionophores on lambs and angora kid goats on rangeland. J. Anim. Sci., 68: 3980-3986.
Direct Link - Kioumarsi, H., K.J. Khorshidi, M. Zahedifar, A.R. Seidavi, S.Z. Mirhosseini and M.R. Taherzadeh, 2008. The effect of dietary energy and protein level on performance, efficiency and carcass characteristics of Taleshi lambs. Asian J. Anim. Vet. Adv., 3: 307-313.
CrossRefDirect Link - Kioumarsi, H., K.J. Khorshidi, M. Zahedifar, A.R. Seidavi, Z.S. Yahaya, W.A. Rahman and S.Z. Mirhosseini, 2008. Estimation of relationships between components of carcass quality and quantity in Taleshi lambs. Asian J. Anim. Vet. Adv., 3: 337-343.
CrossRefDirect Link - Kirk, D.J., L.W. Greene, G.T. Schelling and F.M. Byers, 1985. Effects of monensin on monovalent ion metabolism and tissue concentrations in lambs. J. Anim. Sci., 60: 1479-1484.
Direct Link - Kirk, D.J., L.W. Greene, G.T. Schelling and F.M. Byers, 1985. Effects of monensin on Mg, Ca, P and Zn metabolism and tissue concentrations in lambs. J. Anim. Sci., 60: 1485-1490.
Direct Link - Mbanzamihigo, L., C.J. Van Nevel and D.I. Demeyer, 1996. Lasting effects of monensin on rumen and caecal fermentation in sheep fed a high grain diet. Anim. Feed. Sci. Technol., 62: 215-228.
Direct Link - McGinn, S.M., K.A. Beauchemin, T. Coates and D. Colombatto, 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast and fumaric acid. J. Anim. Sci., 82: 3346-3356.
PubMedDirect Link - Mohamed, O.E., L.D. Satter, R.R. Grummer and F.R. Ehle, 1988. Influence of dietary cottonseed and soybean on milk production and composition. J. Dairy Sci., 71: 2677-2688.
Direct Link - Muwalla, M.M., M.Y. Harb and T.F. Grosby, 1998. Effects of lasolocid and protein levels on the performance of Awassi lambs. Small Rum. Res., 28: 15-22.
Direct Link - Rogers, M., J.P. Jouany, P. Thivend and J.P. Fontenot, 1997. The effects of short-term and long-term monensin supplementation and its subsequent withdrawal on digestion in sheep. Anim. Feed. Sci. Technol., 65: 113-127.
Direct Link - Rowghani, E., M.J. Zamiri and S.R. Ebrahimi, 2006. Effects of monensin and thiamin and their combinations on feedlot performance, blood glucose, BUN levels and carcass characteristics of mehraban lambs fed a high concentrate diet. Pak. J. Biol. Sci., 9: 2835-2840.
CrossRefDirect Link - Rumpler, W.V., D.E. Johnson and D.B. Bates, 1986. The effect of high dietary cation concentration on methanogenesis by steers fed diets with and without ionophores. J. Anim. Sci., 62: 1737-1741.
Direct Link - Russell, J.B., 1987. A proposed mechanism of monensin action in inhibiting ruminal bacterial growth: effects on ion flux and protonmotive force. J. Anim. Sci., 64: 1519-1525.
PubMedDirect Link - Salles, M.S.V., M.A. Zanetti and A.F. Salles, 2008. Effect of monensin on mineral balance in growing ruminants reared under different environmental temperatures. Anim. Feed. Sci. Technol., 141: 233-245.
Direct Link - Schelling, G.T., 1984. Monensin, mode of action in the rumen. J. Anim. Sci., 58: 1518-1527.
Direct Link - Soodeen-Karamath, S. and F.G. Youssef, 1999. Effect of monensin, avoparcin and grass supplementation on utilization of urea-treated rice straw by sheep and goats. Small Rum. Res., 33: 201-211.
Direct Link - Spears, J.W. and R.W. Harvey, 1984. Performance, ruminal and serum characteristics of steers fed lasalocid on pasture. J. Anim. Sci., 58: 460-464.
Direct Link - Staples, C.R., J.M. Burke and W.W. Thatcher, 1998. Influence of supplemental fats on reproductive tissues and performance of lactating cows. J. Dairy Sci., 81: 856-871.
CrossRefPubMedDirect Link - Stock, R.A., S.B. Laudert, W.W. Stroup, E.M. Larson, J.C. Parrott and R.A. Britton, 1995. Effect of monensin and monensin and tylosin combination on feed intake variation of feedlot steers. J. Anim. Sci., 73: 39-44.
Direct Link - Thonney, M.L., E.K. Heide, D.J. Duhaime, R.J. Hand and D.J. Perosio, 1981. Growth, feed efficiency and metabolite concentrations of cattle fed high forage diets with lasalocid or monensin supplements. J. Anim. Sci., 52: 427-433.
Direct Link - Thornton, J.H. and F.N. Owens, 1981. Monensin supplementation and in vivo methane production by steers. J. Anim. Sci., 52: 628-634.
Direct Link - Yang, C.M., C.T. Chang, S.C. Haung and T. Chang, 2003. Effect of lasalocid on growth, blood gases and nutrient utilization in dairy goats fed high forage, low protein diet. J. Dairy. Sci., 86: 3967-3971.
Direct Link - Wallace, R.J., 1994. Ruminal microbiology, biotechnology and ruminant nutrition: Progress and problems. J. Anim. Sci., 72: 2992-3003.
PubMedDirect Link - Webster, A.J.F., 1992. The Metabolizable Protein System for Ruminants. 1st Edn., Department of Animal Husbandry, University of Bristol, Langford, Bristol, UK., pp: 93-110.
Direct Link