
H. Alshiyab
Faculty of Science and Technology, School of Bioscience and Biotechnology, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
M.S. Kalil
Department of Chemical and Process Engineering, Faculty of Engineering, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
A.A. Hamid
Faculty of Science and Technology, School of Bioscience and Biotechnology, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
W.M.W. Yusoff
Faculty of Science and Technology, School of Bioscience and Biotechnology, Universiti Kebangsaan Malaysia, 43600 UKM, Bangi, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 19 | Page No.: 2336-2340







ABSTRACT
The effect of removal of resultant gas resulted in enhancement of the H2 yield. The technique of CO2 scavenging resulted in H2 yield being improved from 408 mL g-1 to reach the maximum of 422 mL g-1. The highest hydrogen productivity of 87.9 ml L-1 h-1 was obtained by CO2 scavenging. Biomass concentration was enhanced to 1.47 g L-1, YP/X of 287 ml g-1 L-1, YX/S of 0.294 and YH2/s of 0.0377 by the use of CO2 scavenging. The results suggested that the presence of the gaseous products in fermentation medium and headspace adversely effect biomass growth and hydrogen production.
PDF Abstract XML References Citation
How to cite this article
H. Alshiyab, M.S. Kalil, A.A. Hamid and W.M.W. Yusoff, 2008. Removal of Headspace CO2 Increases Biological Hydrogen Production by C. acetobutylicum. Pakistan Journal of Biological Sciences, 11: 2336-2340.
DOI: 10.3923/pjbs.2008.2336.2340
URL: https://scialert.net/abstract/?doi=pjbs.2008.2336.2340
DOI: 10.3923/pjbs.2008.2336.2340
URL: https://scialert.net/abstract/?doi=pjbs.2008.2336.2340
INTRODUCTION
Hydrogen has been suggested as a promising alternative to fossil fuels as an energy carrier for transportation needs. Hydrogen gas has a high-energy capacity per unit mass (118.2 kJ g-1) and its reaction with oxygen does not produce greenhouse gases such as CO2. In contrast, methane has a relatively lower energy capacity (36.3 kJ g-1) and CO2 is released during its combustion in conventional engines. However, most hydrogen is usually produced from stored methane reserves and other nonrenewable materials, resulting in net increases of CO2 in the environment. Shifting a fossil fuel economy to a hydrogen economy offers few environmental advantages if both are based on the net consumption of fossil fuels (Wooshin et al., 2005). Thus, it is essential for reducing CO2 emissions that hydrogen production not release a net amount of CO2 into the atmosphere and that the technologies and materials used to produce hydrogen are sustainable.
Major advantages of fermentative H2 production processes are higher evolution rate of H2 and a wide gamut of substrate utilization. However, the inherent disadvantage of these processes is lower yield of H2. This is one of the major deterrents of the fermentative H2 production process. At most a maximum of 4 mol H2 could be obtained per mol glucose during acetate fermentation. To address this problem, efforts are needed to improve the operating conditions to overcome thermodynamic limitations (Fabiano and Perego, 2002) of the acetate fermentation reaction represented below.
C6H12O6+ 2H2O → 2CH3COOH + 2CO2 + 4H2 |
Decrease of H2 partial pressure could be considered as an approach towards improvement of H2 productivity (Levin et al., 2004). Equilibrium constant of the above reaction is according to Le Chatelier`s principle; the equilibrium of the above reaction will shift to the right if one or both of the gaseous products of the reaction is removed. Decrease in partial pressure of H2 by reducing total pressure of the system allows the reaction equilibrium to shift towards right side and thereby enhance the H2 production.
Reducing the partial pressure of hydrogen in reactor vessels can increase hydrogen production. In batch tests, continuous versus intermittent release of gas pressure. Logan et al. (2002) found that continuous release increased hydrogen production by 43%. Reducing the hydrogen partial pressure using a vacuum or by sparging the reactor vessel with nitrogen gas has also increased hydrogen yields (Logan et al., 2002; Tanisho et al., 1998). It was assumed in these tests that this increase in hydrogen production was related to reduction in the hydrogen partial pressure. However, hydrogen can also be consumed via acetogenesis, according to Oh et al. (2003) they mentioned that hydrogen yields were lowered by acetogenesis in the absence of glucose, a heat-treated inoculum produced acetic acid from hydrogen and CO2. The fact that both CO2 and H2 are required for acetogenesis suggested to us that limiting CO2 concentrations in the reactor might provide a viable strategy for reducing acetogenesis. Several studies have shown that produced gases during biohydrogen production can affect hydrogen production. While this increase in production has typically been attributed to a reduced percent of hydrogen in the gas phase, Wooshin et al. (2005) demonstrated that changes in CO2 concentration can also be a factor. They found that overall hydrogen production increased by 43% with chemical scavenging of the CO2 despite an increase in the hydrogen concentration in the headspace. Previous studies by Logan et al. (2002) have shown that an increase in gas pressure in bottles during these tests can reduce hydrogen production. The present study shows that reduction the CO2% of produced gas substantially improves H2 production in an anaerobic fermentation process.
MATERIALS AND METHODS
Microorganism and culture conditions: Clostridium acetobutylicum NCIMB 13357 was purchased from a British culture collection, NCIMB Ltd. Scotland, UK. This study was conducted from 2006 to 2007 in Department of Chemical Engineering; Universiti Kebangsaan Malaysia. The bacterium was cultivated in anaerobic condition in Reinforced Clostridia. Medium (RCM) for 24 h at 30 °C. Liquid medium of RCM was used for inoculum preparation. The growth of culture in RCM was monitored by measuring an optical density at 600 nm using a spectrophotometer. Only inoculum with Optical Density (OD) values greater than 0.4-0.6 after 18 h cultivation was used as inoculum. An inoculum of 10% v/v was used throughout this work. Batch fermentation was carried out at a working volume of 100 in 500 mL rector bottles at 30 °C, each medium was seeded with a 10% inoculum inside anaerobic cabinet and sparged with nitrogen gas (99.9%). All the bottles were tightly closed. The culture pH was not controlled during fermentation and the initial pH was fixed to 7.0 before sterilization process. To quantify H2, gas produced during fermentation was recorded at the end of its production. The evolved gas was collected in a gas collection inverted cylinder and the volume of evolved gas was measured at room temperature by the water displacement method (Morimoto et al., 2004) in a graduated cylinder that had been filled with water of pH 3 or less in order to prevent dissolution of the gas components.
Analytical methods: The gas composition was determined by gas chromatography (Shimadzu Co., Kyoto, GC-8A) under the following conditions: column: Porapack-Q, carrier gas: Nitrogen, flow rate: 33 mL min-1; column temperature: 50 °C, injection temperature: 100 °C, detector temperature: 50 °C, detector: Thermal Conductivity Detector (TCD). The soluble glucose concentration was measured at the end of each batch experiment for the calculation of the amount of glucose consumed by DNS method modified by Miller (1959) using spectrophotometer (UV 1601IPC, Shimadzu Corporation- Japan) optical density (OD550nm). Individual batch experiments were observed until the hydrogen production from each bottle stopped. All bottles (500 mL Scott bottle) with working volume of 100 mL (90% culture medium and 10% inoculum), were initially flushed with nitrogen gas for 5 min and then sealed. CO2 was removed from the headspace using 6 mL of KOH (30% (w/v) (Wooshin et al., 2005) placed in a small tube open only to the headspace. Biotic controls were prepared in the same manner using distilled water instead of KOH.
Gas production was measured using water displacement method (Morimoto et al., 2004) with bio-hydrogen production calculated from headspace gas measurements and the total volume of biogas produced at the end of gas production. The difference in gas production between the treatment and control was used to calculate total CO2 gas reduced by (30% (w/v)) KOH. Hydrogen gas quantity was measured manually from the quantity of the collected gas in inverted cylinder after CO2 absorption by 30% (w/v) KOH. Control was used to calculate the percentage of CO2 absorbed by KOH scavenging method (Wooshin et al., 2005). All of these data were the average (mean) of three trials.
| V1 | = | Total gas volume (Control) |
| V2 | = | Total gas (Scavenging) |
| V3 | = | Collected gas after absorbed by (30% (w/v) KOH) (Control) |
| V4 | = | Collected gas after absorbed by (30% (w/v) KOH) (Scavenging) |
| CO2 (mL) (Scavenging) | = | V1-V2 |
| H2 (mL) (Scavenging) | = | V3-V4 |
To confirm the results gas was collected using plastic bag and analyzed after absorption by 30% (w/v) KOH, by GC- TCD.
RESULTS
It was noted that investigators have reported H2 yield as mol H2 per mol substrate, mol H2 per gram substrate or H2 produced (mL) per gram substrate; hence, for ease of comparison with values reported, the H2 yields were all converted to H2 produced (mL) per gram substrate utilized. The results shown in Table 2 indicated that hydrogen was enhanced 3% by reducing the CO2 concentration in the headspace. Overall, hydrogen yields increased from 408 mL g-1 glucose utilized to 422 mL g-1 glucose utilized with CO2 scavenging. CO2 concentrations in the headspace could not be completely depleted, with CO2 concentrations reduced from 185 mL g-1 (control) to 80 mL g-1 glucose supplied by using the 30% (v/v) KOH trap. As shown in Table 1, trapping the CO2 reduced the measured gas production (compared to that of the control), but increased overall hydrogen production.
| Table 1: | Results of CO2 scavenging by 30% (KOH) on glucose consumption and hydrogen productivity by C. acetobutylicum NCIMB13357. H2 productivity |
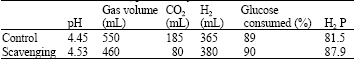 | |
| [Glucose]: 5 g L-1, inoculum size 10% (v/v), initial pH 7.0. Temp. 30 °C | |
| Table 2: | Results of CO2 scavenging by 30% (KOH) on Hydrogen production by C. acetobutylicum NCIMB13357 |
| Y1P/S (H2 mL g-1 glucose supplied) (mL g-1), Y2P/S (mL g-1) (Utilized): (H2 mL g-1 glucose utilized (mL g-1), [Biomass] (g L-1). Biomass production/L culture, YP/X (mL g-1): (H2 mL g-1 Biomass), YX/S: (Biomass production/g glucose supplied), YH2/s (conversion of H2 (mL) to H2 (g) per g glucose utilized) [Glucose]: 5 g L-1, inoculum size 10% (v/v). initial pH. 7.0. Temp 30 °C | |
In the absence of CO2 removal, the pH decrease from pH 7.0 to 4.45 was larger than that of the scavenging route final pH 4.53 due to the higher concentration of CO2 and volatile acids. This slight difference in final pH should not have substantially affected hydrogen production as maximum hydrogen yields in batch tests have been reported to be in the pH range of 5.5-6.0 (Van Ginkel et al., 2001). The results shown in Table 1 indicated that the rate of glucose consumed was increased slightly when CO2 was removed. With CO2 scavenging the bacterial productivity of hydrogen as shown in Table 2 was increased from 81.6 to 87.9 ml L-1 h-1 and that was attributed to the enhancement of biomass concentration due to reduction of CO2% from the produced gas. Reduction of CO2% from the produced gas reduced the pressure of produced gas inside the reactor and finally gave the chance for bacteria for more utilization of substrate to produce gas and to grow and that was clear from the difference between the bacterial growth in both experiments. The results shown in Table 1 demonstrate that bacterial growth was enhanced from 1.38 to 1.47 g L-1 and that was not connected with consumed glucose due to the percentage of glucose consumed mainly same.
A hydrogen concentration of 100% could not be achieved due to incomplete CO2 removal. On the basis of earlier results of Thauer et al. (1977), we would expect that increased hydrogen partial pressures would have inhibited bio-hydrogen production and not resulted in an increase of hydrogen production as observed here. Therefore, the increase in hydrogen production must result from an aspect related to the reduced CO2 concentration in the system. The most likely explanation is that a reduction in CO2 concentrations reduced hydrogen losses via acetogenesis, which could not be completely inhibited. While there was an overall increase in hydrogen production from 408 mL g-1 glucose utilized to 422 mL g-1 glucose utilized with CO2 removal. CO2 could not be completely removed from the headspace. CO2 concentrations may have been disproportionately higher in the liquid phase (relative to equilibrium) where, the CO2 was generated. Thus, it was not possible to completely inhibit acetogenesis due to the remaining CO2 concentration in the liquid phase.
The results shown in Table 1 and 2 suggested that removal the CO2 by chemical scavenging using 30% (KOH) resulted in enhanced the hydrogen yield by 3% and that mainly due to the increased in biomass concentration which was enhanced by reduction of CO2% in produced gas. Different yields were obtained show that the reduction of CO2 from the produced gas reduce the hydrogen production by biomass suggested that the biomass concentration affect inversely on hydrogen production due to byproducts inhibition whereas the biomass per substrate was increased suggested that reduction the gaseous products from the reactor enhanced the growth of biomass.
The results shown in Table 1 demonstrate that CO2 percentage of produced gas was decreased from 34 to 17% (17%) and that was related to the hydrogen percentage in produced gas, which was increased from 66 to 82% (15%). The increase in hydrogen % in produced gas attributed to reduction in CO2% in produced gas and that would reduce the effect of total gas and its pressure inside the reactor. These results were agreed with Wooshin et al. (2005) they reported that by reducing the CO2 concentration in the headspace from 24.5 to 5.2%, hydrogen yield was increased by 43%. They reasoned that to reduction of CO2% in produced gas and its pressure.
The results shown in Table 2 were used to calculate different yields like YP/X (ml g-1 L-1): (H2 ml per g Biomass per L), YX/S: (Biomass production per g glucose supplied) and YH2/s (conversion of H2 (mL) to H2 (g) per g glucose utilized). These results showed that reducing the produced CO2 from headspace that would reduce the percentage of dissolved CO2 in culture medium which showed to enhanced the bacterial growth and finally enhanced the hydrogen productivity as well as hydrogen yield suggested that minimize or removal of produced CO2 from the reactor headspace would enhance the hydrogen production.
DISCUSSION
In general, biological H2 production from organic substrates is limited by the thermodynamics of the hydrogenase reaction, involving the enzyme-catalyzed transfer of electrons from an intracellular electron carrier molecule to protons. On the contrary, protons are poor electron acceptors, so the electron donor must be a strong electron reducing agent. Ferredoxin is a low-potential iron sulfur containing protein that is capable of reducing proton to H2. Another important intracellular electron carrier, NADH, has a higher redox potential. Under actual conditions the ability of reduced ferredoxin and NADH to reduce protons is determined by the redox potential of the overall reaction.
Assuming the intracellular concentrations of the oxidized and reduced form of ferredoxin and NADH are equal, H2 production becomes thermodynamically unfavorable at high H2 partial pressure. This indicates that for ferredoxin, H2 production can continue as long as the partial pressure of H2 is less than ≈ 0.3 atm and for NADH, the partial pressure of H2 must be less than 60 Pa. This implies that at a very low partial pressure (< 60 Pa), NADH could also be used for H2 production. The first attempt to reduce the partial pressure was to use KOH to remove CO2 in the headspace, which decreased the partial pressure of H2 by 87% (Wooshin et al., 2005). This enhanced the yield of H2 from 1.4 to 2 mol H2/mol glucose. Gas sparging is another useful technique to decrease the H2 partial pressure to enhance its yield. Mizuno et al. (2000) observed that the specific H2 production rate increased from 1.5 mL H2/min g biomass to 3.1 mL H2/min g biomass under N2 sparging conditions.
In this study, overall hydrogen production increased here (3%), from 408 to 422 mL g-1 glucose utilized (2.92 to 3.04 mol H2/mol glucose supplied: 3.26 to 3.38 mol H2/mol glucose utilized), with chemical scavenging of the CO2 despite an increase in the hydrogen concentration in the headspace. Previous studies have shown that an increase in gas pressure in reactor vessel can reduce hydrogen production. Present results agreed with reports of Logan et al. (2002) who stated that reducing the pressures of the evolved gases, either by applying vacuum or by gas sparging, could increase hydrogen production. They stated that continuous release of gases increased H2 production by 43% versus tests where there an intermittent release of gas and with Kataoka et al. (1997) who reported that when a vacuum was applied to a reactor operated in chemostat mode, hydrogen production increased by 10%, resulting in an overall yield of 2.2 mol of H2/mol of glucose. For how CO2 affect the bacterial metabolism, only one study was conducted by Ballongue et al. (1987) they reported that the action of CO2 has been studied in yeast in which CO2 acts on enzyme activities level.
An alternate approach to decrease the dissolved hydrogen concentration in a reactor. Tanisho et al. (1998) reported that sparging the reactor with argon gas resulted in an increase in hydrogen yield from 0.52 to 1.58 mol of H2/mol of glucose. In both study, they reasoned the increase of hydrogen production to a reduction in the dissolved hydrogen concentration, the concentration of dissolved CO2 would also have been reduced, contributing to increases in hydrogen production. While applying a vacuum, gas sparging, or CO2 scavenging may all be effective methods of increasing hydrogen production, CO2 scavenging may provide the most direct and cost-effective method of reducing CO2. Gas sparging may also have the undesirable potential to produce a loss of volatile fatty acids in the reactor off gas. It`s known that as the gas pressure increased, the gas would dissolve more in fermentation medium. Finally, there is little information on inhibition by CO2. Certainly most organism appear to be unaffected by CO2 partial pressure up to 0.2 atm. CO2 at high partial pressure will, however, exert an effect through depression of pH or chemical reaction with other compounds in the culture media (Moo-Young, 1985), whereas Doremus et al. (1985) reported that there is no evident that the level of dissolved carbon dioxide affected solvent production by clostridia. Stated that in pure water at 37 °C, Henry`s Low constant, that CO2 would be dissolved 34 times than H2, on molar basis. According to Doremus et al. (1985), the quantity of dissolved CO2 in fermentation medium would affect on bacterial growth and on its metabolism and that was clear in present study and as the produced CO2 reduced by KOH scavenging, the bacterial growth and H2 yield were better than control.
CONCLUSION
The current study, it was only examined the effect of CO2 scavenging on hydrogen production by C. acetobutylicum NCIMB 13357 in batch tests. Even in these tests, it was not possible to completely scavenge CO2 from the headspace during high rates of hydrogen production. If hydrogen is produced in continuous culture, it is not known how effectively it could scavenge CO2 under these conditions. Thus, additional research is needed to extend the batch test results from this test to continuous culture tests, particularly under conditions that affect the hydrogen production rate, such as under different organic loadings and reactor detention times.
REFERENCES
- Ballongue, J., E. Masion, J. Amine, H. Petitdemange and R. Gay, 1987. Inhibitor effect of products of metabolism on growth of Clostridium acetobutylicum. Applied Microbial. Biotechnol., 26: 568-573.
Direct Link - Collet, C., N. Adler, J.P. Schwitzguebel and P. Peringer, 2004. Hydrogen production by Clostridium thermolacticum during continuous fermentation of lactose. Int. J. Hydrogen Energy, 29: 1479-1485.
CrossRef - Doremus, M.G., J.C. Linden and A.R. Moreira, 1985. Agitation and pressure effect on acetone butanol fermentation. Biotech. Bioeng., 27: 852-860.
Direct Link - Kataoka, N., A. Miya and K. Kiriyama, 1997. Studies on hydrogen production by continuous culture system of hydrogen producing anaerobic bacteria. Water Sci. Technol., 36: 41-47.
Direct Link - Logan, B.E., S.E. Oh, I.S. Kim and S. Van Ginkel, 2002. Biological hydrogen production measured in batch anaerobic respirometer. Environ. Sci. Technol., 36: 2530-2535.
CrossRefDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Morimoto, M., M. Atsuko, A.A.Y. Atif, M.A. Ngan and A. Fakhrul-Razi et al., 2004. Biological production of hydrogen from glucose by natural anaerobic microflora. Int. J. Hydrogen Energy, 29: 709-713.
CrossRefDirect Link - Oh, Y.K., E.H. Seol, J.R. Kim and S. Park, 2003. Fermentative biohydrogen production by a new chemoheterotrophic bacterium Citrobacter sp. Y19. Int. J. Hydrogen Energy, 28: 1353-1359.
CrossRefDirect Link - Tanisho, S., M. Kuromoto and N. Kadokura, 1998. Effect of CO2 removal on hydrogen production by fermentation. Int. J. Hydrogen Energy, 23: 559-563.
CrossRefDirect Link - Thauer, R.K., K. Jungerman and K. Decker, 1977. Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev., 41: 100-180.
Direct Link - Van Ginkel, S., S. Sung and J.J. Lay, 2001. Biohydrogen production as a function of pH and substrate concentration. Environ. Sci. Technol., 35: 4726-4730.
CrossRefDirect Link - Yokoi, H., T. Tokushige, J. Hirose, S. Hayashi and Y. Takasaki, 1998. H2 production from starch by a mixed culture of Clostridium butyricum and Enterobacter aerogenes. Biotechnol. Lett., 20: 143-147.
CrossRefDirect Link - Fabiano, B. and P. Perego, 2002. Thermodynamic study and optimization of hydrogen production by Enterobacter aerogenes. Int. J. Hydrogen Energy, 27: 149-156.
CrossRefDirect Link - Levin, D., L. Pitt and M. Love, 2004. Biohydrogen production: Prospects and limitations to practical application. Int. J. Hydrogen Energy, 29: 173-185.
CrossRefDirect Link