
Y. Nikpour
Khoramshahr University of Marine Science and Technology, Iran
H. Zolgharnein
Khoramshahr University of Marine Science and Technology, Iran
M. Sinaei
Khoramshahr University of Marine Science and Technology, Iran
H. Najafzadeh
Chamran University, Ahvaz, Iran
M. Ghavasi
Khoramshahr University of Marine Science and Technology, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 18 | Page No.: 2269-2273







ABSTRACT
The effect of mercury exposure to total Metallothionein (MT) response and bioaccumulation under control and acute mercury exposure were investigated in scats (Scatophagus argus). Scats were exposed to different mercury concentrations (10, 20 and 30 μg Hg L-1) for 24, 48 and 72 h. Total MT levels were determined by Enzyme-Linked Immunosorbent Assay (ELISA) method. Mercury contents were determined through cold Vapour atomic Absorption spectrometry. Induction of MT during exposure was tissue specific, displaying different response pattern in gill and liver. Mercury accumulated much stronger in liver than gill and the latter also showed lower MT level. MT biosynthesis in liver showed a significant increase after exposure to different mercury concentration during different times. This increase was significantly correlated with mercury bioaccumulation. In contrast, presence of different mercury concentration during different times did not significantly modify total MT except for 72 h exposure at 30 μg L-1 in gills. The results suggest that this form of MT existing in S. argus was Hg-inducible and could be extended the as a biomarker of mercury pollution in ecosystems.
PDF Abstract XML References Citation
How to cite this article
Y. Nikpour, H. Zolgharnein, M. Sinaei, H. Najafzadeh and M. Ghavasi, 2008. Evaluation of Metallothionein Expression as a Biomarker of Mercury Exposure in Scatophagus argus. Pakistan Journal of Biological Sciences, 11: 2269-2273.
DOI: 10.3923/pjbs.2008.2269.2273
URL: https://scialert.net/abstract/?doi=pjbs.2008.2269.2273
DOI: 10.3923/pjbs.2008.2269.2273
URL: https://scialert.net/abstract/?doi=pjbs.2008.2269.2273
INTRODUCTION
Marine ecosystems are contaminated by different pollutants especially metals due to human activities. Mercury pollution is one of the world,s most serious environmental problems (Pilon-Smith and Pilon, 2000). Mercury exposure is the second most common cause of toxic metal poisoning. It exists in both organic and inorganic forms and is among the few pollutants that exhibit biomagnification in aquatic food chain (Ribeiro et al., 1996). Public health concern over mercury exposure due to contamination of fish with methyl mercury has long been a topic of debate. Existing situation in the environment has prompted numerous investigations to consider effect of mercury on the biological functions of marine organisms, particularly defense mechanism in fish. Different fish species show differences in metal accumulation and metallothionein biosynthesis as a detoxification mechanism. Tissue also accumulates metals to a different extend due to the differences in physiological and biochemical functions.
The distributions of scats in the pacific region are in harbors, natural embayments, estuaries and lower parts of freshwater streams, frequently among mangroves. This species often exist in small aggregation.
Metallothionein among a class of proteins with relatively low molecular weight 6-8 (kDa) characterized by the intrinsic presence of 20 cysteine group in their structure which confers unique metal binding properties to the molecules (Dabrio et al., 2002; Chan et al., 2002). MT occurs mainly in the cytosol and is also present in the nucleus (Decataldo et al., 2004). Metallothioneins involves in sequestration of toxic (Cd, Hg) and essential (Cu, Zn) metals have been proposed as sensitive biomarker in assessing metal exposure and prediction of potential detrimental effects induced by metal contamination (Ivankovic et al., 2005 ). The induction of MT or similar metal binding proteins in fish was demonstrated either under experimental or environmental conditions (De Boeck et al., 2003; Hamza-Chaffai et al., 2000).
In the present study, S. argus were exposed to different levels of mercury for different times. Bioaccumulation of Hg and MT biosynthesis in tissues (gill and liver) were measured in such experimental series at the end of each exposure period. The aim of this study was to assess Hg-binding capacity of two organs (liver and gill) to biosynthesis metallothionein, to examining the relationship between MT and Hg concentrations for each tissue and to determining effects of time and dose on MT response in these tissues.
MATERIALS AND METHODS
Fish holding condition: Fishes (S. argus) were collected from west coast of Persain gulf during summer 2007. Male and Female scats (with a mean weight of 143±7 g and total length 14±1 cm) were transferred to laboratory of Khoramshahr Marine Science and Technology University (KMSU, Iran). They were maintained in two 200 L aquaria for 2 weeks before initiation of the exposure study. They were acclimatized to aerated and standard OECD water (Organization for Economic Cooperation and Development, 1993) at temperature of 26±1°C under normal photoperiod of 12-14 h. The water Hardness was 250 mg L-1 as CaCO3 and pH 7.4±0.02. The medium used was filtered and the levels of NH4, NO2 and NO3 in the water recorded to be within 0.1, 0.1 and 20 mg L-1, respectively. During acclimatization, scats were being fed once a day with Biomar Co. fish food at a rate of 1% of fish biomass.
Waterborne mercury exposure experiment: Test scats (n = 5 for each concentration) were exposed to three contamination levels 0, 10, 20 and 30 μg L-1 for 24, 48 and 72 h at 26±1°C exposure duration by addition of mercury from a mercury stock solution prepared in deionised water (HgCl2, 20, extra pure, merck). No mortality were observed during the experiment . Water replacement was carried out every day and the mercury concentrations in the water were determined by CVAAS. The ambient hardness was 250 mg L-1 as CaCO3 and the water pH varied between 7.37 to 7.68. Scats were not fed during the experiment and were kept starved for 24 h before the experiment. After each exposure period Scats were anesthetized by dry extract of clove pink, then liver and gill samples were dissected on ice. The samples were divided into two parts, weighed and stored at -80°C for further processing.
Mercury analysis: Total mercury level were determined using cold vapour analysis technique. After thawing, 1 g of tissue was digested in 20 mL of 3:1 concentrated redistilled HNO3 and concentrated H2SO4, and then oxidized with 10 mL of saturated solution of KMnO4. Excess oxidizing agents and mercury ions were reduced by 10 mL of reducing solution (3% NaBH4 in 1% NaOH) in a hydride generator apparatus, and thereafter mercury was vaporized and measured in the atomic absorption spectrophotometer (Unicam 919). The instrument was pre-calibrated using standard solutions prepared from commercial Hg chemical of analytical grade. A blank (n = 3) was run in the same manner as that of samples and mercury was determined using standard prepared in the same acid matrix which did not show significant metal contamination.
MT analysis: After thawing, the gill and liver samples were prepared by individually homogenizing in homogenization buffer (10 mM cold Tris-HCl pH 7.0) containing 5 m M2-mercaptoethanol to prevent oxidation with phenylmethanesulfonylfluoride (PMSF, protease inhibitor) in a 1:2.5-3.0 (w/v) volume using a Teflon homogenizer at 1000-1200 rpm. The homogenates were centrifuged at 12000 x g for 40 min at 4°C. The supernatant was heated at 80°C for 10 min in order to denature thermolabile proteins, and then centrifuged again at 12000 x g for 40 min at 4°C.
Ninety six well palates were coated with 100 μL of the different samples for 12 h at 4°C. The saturation was realized for 2 h at ambient temperature with 200 μL of a 3% Bovine Serum Albumin (BSA) in 0.01 mol L-1 Phosphate Buffered Serum (PBS) albumin at pH 7.4. After 4 rinses with 0.01% BSA, 0.05% Tween 20 in PBS, 10 μL of polyclonal antibody (Rabbit Anti-cod metallothionein diluted 1:1000) were added to each well and incubated for 2 h at 37°C . After four rinses with previous buffer, 100 μL of HRP (Peroxidase labeled goat anti-Rabbit IgG) diluted 1:3000 in TBS-Tween was added and incubated for 2 h. After 4 washes, 100 μL well-1 of the ABTS peroxidase substrate (Kirkegaard and Perry Lab, USA) was added followed by incubation at room temperature for 20-30 min. Colour development was measured at 405 nm with an automatic micro-titer plate ELISA reader. The liner regression coefficient (Microsoft Excel 97 SR-1, 1997, Microsoft Corp, Seattle WA.USA) for the logarithm for the MT standard concentrations was -0.99 and the slope was -0.2.
Statistical analysis: Statistical analysis of data was carried out using SPSS Statistical Package Programs (version 13). Data were tested for homogeneity of variance and normal distribution. ANOVA was calculated. A post host comparison was made using Tukey,s tests. Differences between means were testes at 5% probability level. Diagrams were drawn using Microsoft Excel.
RESULTS AND DISCUSSION
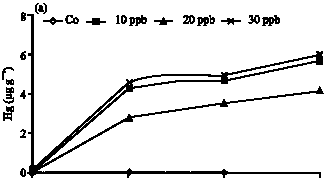
Mercury bioaccumulation: Mercury concentration in the liver was significantly higher than gills. Mercury bioaccumulation appeared to be strongly correlated with contamination level of water and exposure duration (p>0.05). However no interaction was found between these two factors.
Mercury bioaccumulation was rapid during 24 h exposure in the liver then a plateau tendency was seen between 24 to 48 h in 10 μg Hg L-1 level and thereafter gradual and slow in 20 and 30 μg Hg L-1 level at 48 h exposure. Mercury bioaccumulation at 30 μg Hg L-1 24 h exposure was on the other hand lower than exposure in 20 μg Hg L-1 even after 72 h exposure (Fig. 1a).
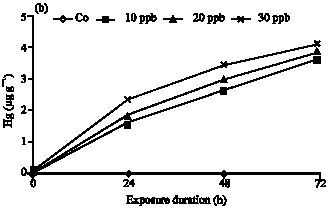
| Fig. 1: | Mercury bioaccumulation at the (a) liver and (b) gill of S. argus as a function of three levels of contamination in the water column (Cw) and of the exposure duration (Ed) |
| Table 1: | Mean concentration (±SD) of MTs in liver and gill of scats; unexposed and exposed to Hg |
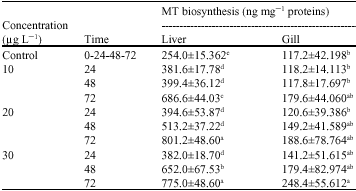 | |
The values followed by the same letter are not statistically different among the treatment | |
In contrast, the accumulation rate showed a significant increases in gills with time close to linearity but bioaccumulation at 24 h exposure in 20 and 30 μg Hg L-1 levels was lower than 72 h exposure in 10 and 20 μg Hg L-1 (Fig. 1b).
MT biosynthesis: MT biosynthesis strongly occurred in the tissues showing significantly higher level in liver than gill (Fig. 2). The results further indicated significant differences between control and treatments in the liver. A two way ANOVA (contamination level and exposure duration) performed on MT concentrations was also significant (p<0.05), indicating the effect of contamination levels of mercury and effect of exposure duration on MT biosynthesis. The interaction between contamination and Exposure duration was also significant (p<0.05).
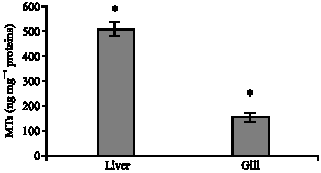 | |
| Fig. 2: | MT biosynthesis in two organ level (liver and gill) of S. argus exposed to Hg. *Significant at p<0.05 |
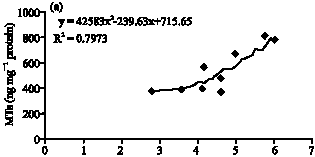
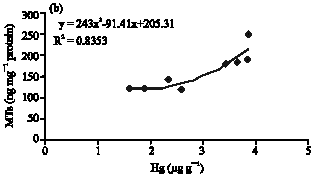
| Fig. 3: | Correlation between MT concentration and Hg bioaccumulation in the (a) liver and (B) gill of S. argus for 72 h to Hg. R2 = 0.46 are significant |
The data envisage no significant differences between 24 and 48 h exposure time at 10 μg Hg L-1 and also the two higher contamination level for 24 h exposure duration in liver (Table 1). It is manifested from the data that the effect of exposure duration and contamination level was significant but no inter-relationship was found in the gill. On the other hand no significant differences was established between control and treatment excepting at the higher level and long exposure duration in the gill (30 μg Hg L-1 72 h). Among treatments 10 μg Hg L-1 for 24 and 48 h and 20 μg Hg L-1 for 24 h differ significantly with 30 μg Hg L-1 for 72 h in gill (Table 1) while no differences was found in other treatments irrespective of exposure duration.
Correlation between metal accumulation and the corresponding MT level in S. argus: Increase in MT biosynthesis in the liver of scats for 72 h exposure to mercury levels were highly correlated (R2 = 0.79, p = 0.001) with the increases mercury concentration (Fig. 3a). Although no significant difference was found in the gills between control and exposed scat except for 30 μg L-1 after 72 h, high correlation (R2 = 0.83, p = 0.001) between MT and mercury was found (Fig. 3b).
DISCUSSION
Short term exposure of scat to different mercury concentrations ranging from 0 to 30 μg L-1 during different period of exposure resulted in increase in mercury accumulation in the gill and liver. Mercury was accumulated significantly in the liver compared to gill. This accumulation order was also supported by several authors and might be attributed to the lower metal-binding capacity of the gills as a consequence of the low gill MT concentrations present (Cattani et al., 1996; De Smet and Blust, 2001; Lange et al., 2002). Olsvik et al. (2001) suggested that the cadmium present in the gills of trout is rapidly cleaned to the circulation system to the liver and kidney where it could be retained for a longer time.
MT contents of these tissues appeared to be exposure and level of metal dependent during 24-72 h period. Elsewhere it has been shown that the MT content appears to have time and dose dependent in vivo and in vitro studies. Wu and Hwang (2005) also corroborated this findings in the liver and gill of Tilapia (Oreochromis mosambicus) after exposure to cadmium during 24-72 h. This result implies that dose-related response of MT expression only occur with doses of heavy metals that do not cause detrimental effects to the physiological functioning of the fish (Wu et al., 2002). Correlation among MT expression, heavy metal accumulation, and tolerance of fish to heavy metals are complicated, specially in in vivo systems (Wu et al., 2002) similar changed appeared in this in vitro study.
The present results clearly indicate tissue -specific differences of MT induction in response to mercury exposure. Liver can induce MT biosynthesis much higher than gill. Gill MT level showed only a moderate increase during the exposure experiment. Indeed no significant mercury related induction of gill MT could be detected in the scat even at the highest exposure level and duration reflecting apparently low capacity of the gill for Hg-induction after a short water-borne mercury. Chaffai et al. (1997) and Olsvik et al. (2001) have suggested that the gill do not constitute a good organ for MT quantification perhaps because MT induction is dependent on the cell type and occur primarily in the chloride cell (Burkhardt-Holm et al., 1999; Dang et al., 2000). But this result shows that a clear correlation exists between mercury and MT level in the gill. It is remarkable that scat which show the best survival rate under mercury exposure has the fastest and target organ during exposure. Mercury toxicity in fish shows a typical shock phase with the extensive damage in the first hours or days of exposure and repair thereafter (McDonald and Wood, 1993) and a fast protective response is thus a clear advantage.
This data also indicated a good correlation between Hg and MT levels in the liver of scat. This positive correlation have also been observed in roach liver exposed to Cd (Bonwick et al., 1991) and in common carp and gibel carp (De Boeck et al., 2003), as well as gudgeon exposed to increased Zn concentrations in the field (Bervoets et al., 2002). Filipovic and Raspor (2003) have interpreted the positive correlations between metal and metallothionein content in fish tissue as the metal sequestration by MT and a poor correlation as metal exceeding the binding capacity of MT or the involvement of non-MT proteins. According to this interpretation metal binding capacity of liver (Table 1). Mean concentration (±SD) of MTs in liver and gill of scats is unexposed and exposed to Hg. The values followed by the same letter are not statistically different among the treatment is not exceeded at any mercury concentration and exposure duration measured in this study and provides the mechanism for the high mercury bioaccumulation capacity of this organ.
The results from this study show that liver is more efficient organ than gill to induce MT biosynthesis. There is a significant and early increase of MT biosynthesis in S. argus after exposure to Hg in liver but in gill only after higher contamination level occurs. Results also show significant effect of contamination level and exposure duration on MT concentration in S. argus. The increase was significantly correlated to Hg bioaccumulation. The advantage in using scat is the rapid response within 24 h. These parameter could be extend for use of MT as a biomarker of mercury pollution in marine ecosystems in S. argus.
ACKNOWLEDGMENT
Thanks are due to Dr. A.N Mohanty (CIFA) Bhubaneswer India for critically going through the manuscript and suggesting improvement.
REFERENCES
- Bervoets, L., M. Lodts, K. Van Campenhout and R. Blust, 2002. Heavy metals as a threat for restored fish populations in a lowland river. In: Conservation of Freshwater Fishes: Options for the Future. Collares-Pereira, M.J., M.M. Coelho and I.G. Coux (Eds.). Blackwell Sci. Publ., Oxford, ISBN-10: 0852382863, pp: 250-261.
Direct Link - Bonwick, G.A., P.R. Fielden and D.H. Davies, 1991. Hepatic metallothionein levels in roach (Rutilus rutilus) continuously exposed to water-borne cadmium. Comp. Biochem. Physiol., 99C: 119-125.
Direct Link - Cattani, O., R. Serra, G. Isani, R. Giampaolo, P. Cortesi and E. Carpene, 1996. Correlation between metallothionein and energy metabolism in sea bass, Dicentrarchus labrax, exposed to cadmium. Comp. Biochem. Physiol., 113C: 193-199.
CrossRefDirect Link - Dabrio, M., A.R. Rodriguez, G. Bordin, M.J. Bebianno, M. De Leya and et al., 2002. Recent developments in quantification methods for metallothionein. J. Inorg. Biochem., 88: 123-134.
Direct Link - De Smet, H. and R. Blust, 2001. Stress responses and changes in protein metabolism in carp Cyprinus carpio during cadmium exposure. Ecotoxicol. Environ. Safety, 48: 255-262.
Direct Link - De Boeck, G., T.T.H. Ngo, K. Van Campenhout and R. Blust, 2003. Differential metallothionein induction patterns in three freshwater fish during sublethal copper exposure. Aquat. Toxicol., 65: 413-424.
Direct Link - Decataldo, A., A. Di Leo, S. Giandomenico and N. Cardellicchio, 2004. Association of metals (mercury, cadmium and zinc) with metallothionein-like proteins in storage organs of stranded dolphins from the Mediterranean sea (Southern Italy). J. Environ. Monit., 6: 361-367.
Direct Link - Hamza-Chaffai, A., J.C. Amiard, J. Pellerin, L. Joux and B. Berthet, 2000. The potential use of metallothionein in the clam Ruditapes decussatusas a biomarker of in situ metal exposure. Comp. Biochem. Physiol., C127: 185-197.
PubMedDirect Link - Ivankovic, D., J. Pavicic, M. Erk, V. Filipovic-Marijic and B. Raspor, 2005. Evaluation of the Mytilus galloprovincialis Lam. digestive gland metallothionein as a biomarker in a long-term field study: Seasonal and spatial variability. Mar. Pollut. Bull., 50: 1303-1313.
Direct Link - Olsvik, P.A., K. Hindar, K.E. Zachariassen and R.A Andersen, 2001. Brown trout (Salma trutta) metallothioneins as biomarkers for metal exposure in two Norwegian rivers. Biomarkers.,q 6: 274-288.
CrossRef - Ribeiro, C.A.O., J.R.D. Guimara˜es and W.C. Pfeiffer, 1996. Accumulation and distribution of inorganic mercury in a tropical fish (Trichomycterus zonatus). Ecotox. Environ. Safety., 34: 190-195.
Direct Link - Wu, S.M. and P.P. Hwang, 2005. Copper or cadmium pretreatment increases the protection against cadmium toxicity in tilapia larvae (Oreochromis mossambicus). Zool. Studies., 42: 179-185.
Direct Link