
H. Alshiyab
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
M.S. Kalil
Department of Chemical and Process Engineering, Faculty of Engineering, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
A.A. Hamid
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
W.M. Wan Yusoff
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 UKM Bangi, Selangor, Malaysia
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 18 | Page No.: 2193-2200







ABSTRACT
The objective of this study is to investigate the effect of salts addition to fermentation medium on hydrogen production, under anaerobic batch culture system. In this study, batch experiments were conducted to investigate the inhibitory effect of both NaCl and sodium acetate on hydrogen production. The optimum pH and temperature for hydrogen production were at initial pH of 7.0 and 30°C. Enhanced production of hydrogen, using glucose as substrate was achieved. In the absence of Sodium Chloride and Sodium Acetate enhanced hydrogen yield (YP/S) from 350 mL g-1 glucose utilized to 391 mL g-1 glucose utilized with maximum hydrogen productivity of 77.5 ml/L/h. Results also show that sodium chloride and sodium acetate in the medium adversely affect growth. Hydrogen yield per biomass (YP/X) of 254 ml/L/g, biomass per substrate utilized (YX/S) of 0.268 and (YH2/S) of 0.0349. The results suggested that Sodium at any concentration resulted to inhibit the bacterial productivity of hydrogen.
PDF Abstract XML References Citation
How to cite this article
H. Alshiyab, M.S. Kalil, A.A. Hamid and W.M. Wan Yusoff, 2008. Effect of Salts Addition on Hydrogen Production by C. acetobutylicum. Pakistan Journal of Biological Sciences, 11: 2193-2200.
DOI: 10.3923/pjbs.2008.2193.2200
URL: https://scialert.net/abstract/?doi=pjbs.2008.2193.2200
DOI: 10.3923/pjbs.2008.2193.2200
URL: https://scialert.net/abstract/?doi=pjbs.2008.2193.2200
INTRODUCTION
The modern economy is largely built on combustion of fossil fuels such as oil, coal and natural gas. This will eventually lead to the depletion of limited fossil energy resource. Furthermore, the combustion of fossil energy source contributed to heavy air pollution and the greenhouse effect. To reduce air pollution and greenhouse gas emissions, H2 has been considered to be an ideal energy alternative for the future. Compared with conventional H2 generation processes, biological H2 production using fermentative and photosynthetic bacteria is not only environmental friendly, but also energy saving (Das and Veziroglu, 2001).
Biohydrogen fermentation by anaerobic fermentation by anaerobic process is greatly influenced by many factors, such as pH, temperature, toxic substance and ionic strength (Zheng et al., 2005).
The effects of the different factors, such as substrate, bacteria strain, reactors used and conditions on anaerobic fermentation for hydrogen production have been investigated. However few study investigated the impact of metal ion concentration on bio-hydrogen production. Lee et al. (2001) only studied the effect of normal metal ion iron concentration in the external environment on hydrogen fermentation using FeC12, treating sucrose solution by the mixed microorganisms from a soybean-meal silo and found out the relationship between iron concentration and VFAs and solvents. Subsequently, Lin and Lay (2005) reported that magnesium, sodium, zinc and iron were important trace metals affecting hydrogen production magnesium being the most significant. Metal ions are easily found in wastewaters, whereas in practice some amount of sodium salts is frequently found in various wastewaters. The presence of high sodium ion concentration is considered to have an inhibitory effect on the anaerobic wastewater treatment and activated sludge treatment (Xiaolong et al., 2006).
Earlier investigations on sodium inhibition were mainly concentrated on undomesticated methanogenic bacteria in anaerobic digestion process; and under different conditions, different bacteria (microbes) have different tolerances to sodium toxicity. However, few study have been done on the effect of sodium salt on hydrogen-producing bacteria (pure bacteria). However, ionic strength is one of the important factors influencing H2 production because the growth of fermentative bacteria requires a number of ions such as sodium and potassium. The ionic strength in the solution is determined by both ion species and its concentrations and it can be either stimulatory or inhibitory and even toxic to H2 production. A low level of ionic strength may have no or even stimulatory effect on H2 production. However, a high level of ionic strength can result in cell lysis (Niel et al., 2003). Therefore, a proper ionic strength is necessary to optimize anaerobic hydrogen production process.
Inhibitory effect of ionic strength on hydrogen fermentation is available in the literature. Therefore this study was conducted to investigate the influence of ionic strength on glucose degradation, hydrogen production. The ionic strength in the solution was provided in the form of sodium chloride and sodium acetate. New medium we formulated in the lab for hydrogen production using C. acetobutylicum NCIMB 13357 and glucose is the sole carbon source.
MATERIALS AND METHODS
Microorganism and culture conditions: C. acetobutylicum NCIMB 13357 was purchased from a British culture collection, NCIMB Ltd. Scotland, UK. This study was conducted from 2006 to 2007 in Department of Chemical Engineering; Universiti Kebangsaan Malaysia. The bacterium was cultivated in anaerobic condition in Reinforced Clostridial Medium (RCM) for 24 h at 30 °C. Liquid medium of RCM was used for inoculum preparation. The growth of culture in RCM was monitored by measuring an optical density at 600 nm using a spectrophotometer. Only inoculum with optical density (OD) values greater than 0.4 after 18 h cultivation was used as inoculum. An inoculum of 10% (v/v) was used throughout this study.
Cultivation medium: New medium was formulated in the lab to be used for hydrogen production have the following composition in (g L-1): glucose (5), yeast extract (5), L-cystine. HCl (1.0) and bacteriological agar (0.5). The initial anaerobic condition in the reactor after inoculation inside the anaerobic glove box was established by replacing the gaseous phase with nitrogen at start of cultivation. Then incubated at 30°C in temperature controlled water bath without shaking. The evolved gas was monitored and collected in a gas collection cylinder and the volume of evolved gas was measured at room temperature by the water displacement method (Morimoto et al., 2004) in a graduated cylinder (inverted), that had been filled with water of pH 3 or less in order to prevent dissolution of the gas components.
Analytical methods: The gas composition was determined by gas chromatography (Shimadzu Co., Kyoto, GC-8A) under the following conditions: column: Porapack-Q, carrier gas: Nitrogen, flow rate: 33 mL min-1; column temperature: 50°C; injection temperature: 100°C; detector temperature: 50°C, detector: Thermal Conductivity Detector (TCD). The soluble glucose concentration was measured at the end of each batch experiment for the calculation of the amount of glucose consumed by DNS method modified by Miller, (1959) using spectrophotometer (UV 1601IPC, Shimadzu corporation- Japan) optical density (OD550 nm). Individual batch experiments were observed until the hydrogen production from each bottle stopped. Final medium pH was measured by pH meter (Mettler Toledo) and final biomass was measured by Spectrophotometer (UV 1601IPC, Shimadzu corporation- Japan) at optical density (OD600 nm). All of these data were the average (mean) of three trials.
Experimental design: Batch experiments were conducted with 500-mL Scotch bottle with working volume of 100 mL (90% medium and 10% inoculum). Different concentrations of NaCl and NaCH3COOH were used to find out its effect on fermentative bacteria we used in this study, ranging from 0.0 to 5000 mg L-1 for NaCl and from 0.0 to 3000 mg L-1 for NaCH3COOH.
RESULTS AND DISCUSSION
The results shown in Fig. 1 and 5 indicated that at 0.0 g L-1 of NaCl or NaCH3COOH, the maximum H2 yield was obtained at 0.0 g L-1 of NaCl was 391 mL g-1 glucose utilized and for NaCH3COOH was 355 mL g-1 glucose utilized. The increase of hydrogen yield was reversed to bacterial productivity of hydrogen which resulted to increase as shown in Fig. 3 and 6 which indicated that hydrogen productivity was dropped from 77.5 ml/L/h at 0.0 g L-1 for NaCl to 63.5 ml/L/h at 5 g L-1 and for NaCH3COOH from 72.5 ml/L/h at 0.0 g L-1 to 66.5 ml/L/h at 3 g L-1. The maximum productivity of 77.5 ml/L/h obtained by using this medium was lower than 137 ml/L/h which reported by Morimoto et al. (2004) at 50°C using POME sludge and the other reported values was for continuous process not batch culture.
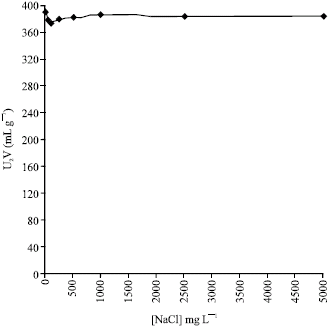 | |
| Fig. 1: | Effect of NaCl addition on H2 yield (mL g-1) utilized |
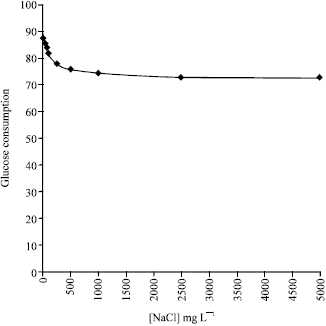 | |
| Fig. 2: | Effect of NaCl addition on glucose consumption (%) |
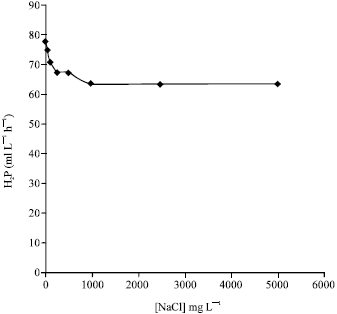 | |
| Fig. 3: | Effect of NaCl addition on Hydrogen productivity H2P (ml/L/h) |
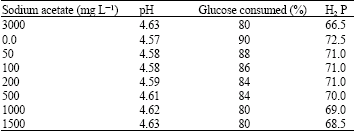
The glucose consumption was influenced by the presence of NaCl or NaCH3COOH or both in fermentation medium. The results shown in Fig. 2 and 6 indicated that with increase in NaCl from 0.0 to 5000 mg L-1 and NaCH3COOH from 0.0 to 3000 mg L-1, the glucose consumption decreased gradually. For the control, both NaCl (5 g L-1) was 73% and NaCH3COOH (3 g L-1) was 80%. Compared with the control, at 0.0 mg L-1 of NaCl and NaCH3COOH, the glucose consumption was enhanced and reached to 87 and 90%, respectively, but in the presence of a low level of NaCl or NaCH3COOH, glucose consumption was decreased as the concentration of each was increased. This demonstrates that glucose consumption by C. acetobutylicum NCIMB13357 was influenced by the presence of any amount of NaCl or NaCH3COOH in the fermentation medium and that might be the inhibitory effect of sodium at any concentration. The results showed that, NaCl and NaCH3COOH affect in negative way on glucose degradation.
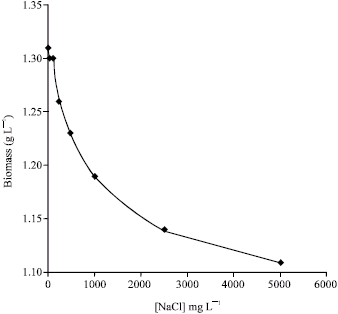 | |
| Fig. 4: | Effect of NaCl addition on Biomass concentration (g L-1) Glucose 0.5 g L-1, inoculum size 10% (v/v), I pH 7.0, Temp. 30°C |
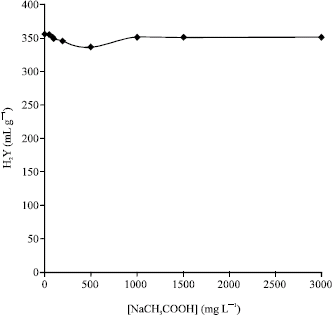 | |
| Fig. 5: | Effect of NaCH3COOH addition H2 yield (mL g-1) Utilized |
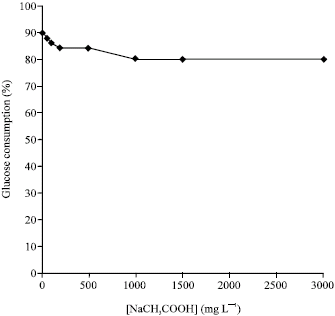 | |
| Fig. 6: | Effect of NaCH3COOH addition on glucose consumption (%) |
| Table 1: | Results the effect of NaCl on glucose consumption and H2P (ml/L/h) by C. acetobutylicum NCIMB13357 |
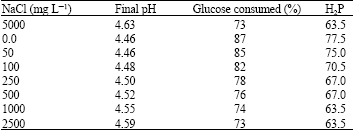 | |
| Glucose: 5 g L-1, inoculum size 10% (v/v) I pH 7.0. Temp. 30 °C | |
| Table 2: | Results the effect of NaCl on hydrogen production by C. acetobutylicum NCIMB13357 |
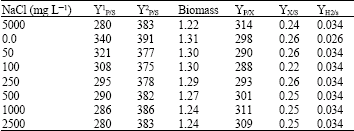 | |
| Y1P/S (H2 mL g-1 glucose supplied) (mL g-1)), Y2P/S (mL g-1) (Utilized): (H2 mL g-1 glucose utilized), [Biomass] (g L-1). Biomass production g L-1 culture, YP/X (mL/g/L): (H2 mL g-1 Biomass L-1), YX/S: (Biomass production per g glucose supplied), YH2/s: (conversion of H2 mL to H2 (g) per g glucose utilized) Glucose: 5 g L-1, inoculum size 10% (v/v), I pH 7.0, Temp. 30 °C | |
Obtained data shown in Table 1 and 2 demonstrated that hydrogen production depends on the presence of NaCl and its concentration and as NaCl concentration increased the produced hydrogen was decreased. Niel et al. (2003) reasoned that inhibition of decreased hydrogen production to increase in concentration of the NaCl in the fermentation medium and suggested that due to the increase of the ionic strength in the fermentation medium concluded that the high ionic strength affect the bacterium metabolic pathway.
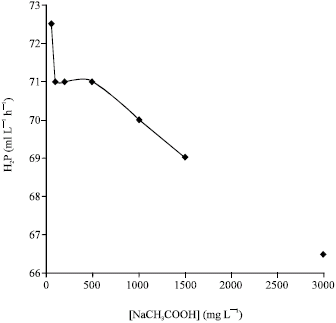 | |
| Fig. 7: | Effect of NaCH3COOH addition on hydrogen productivity H2P (ml/L/h) |
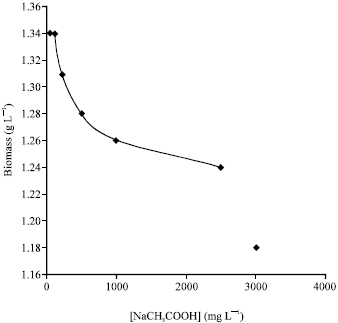 | |
| Fig. 8: | Effect of NaCH3COOH addition on biomass concentration (g L-1) glucose 0.5 g L-1, inoculum size 10% (v/v), I pH 7.0, Temp. 30°C |
The effect of NaCl it was also shown on the growth of bacteria. It appeared that the concentration of inhibitor affects the bacterial growth and as concentration increased, the final biomass concentration was lower. The results shown in Fig. 4 and 8 shown that the presence of NaCl or NaCH3COOH in fermentation medium affect on bacterial growth with maximum biomass concentration was enhanced from 1.22 at 5 to 1.31 at 0.0 g L-1 whereas for NaCH3COOH enhanced from 1.18 at 3 to 1.34 at 0.0 g L-1, suggested that both components were inhibited the bacterial growth and that resulted to reduce the produced hydrogen. Niel et al. (2003) suggested that high salt concentration affect the bacterial growth and resulted to cell lyses.
| Table 3: | Results the effect of sodium Acetate (NaCH3COOH), on glucose consumption and H2 P (ml/L/h) by C. acetobutylicum NCIMB13357 |
 | |
| Glucose: 5 g L-1, inoculum size 10% (v/v) I pH 7.0. Temp 30 °C | |
| Table 4: | Results the effect of sodium acetate (NaCH3COOH), on hydrogen production by C. acetobutylicum NCIMB13357 |
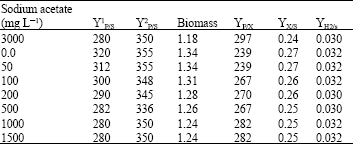 | |
| Y1P/S(H2 mL g-1 glucose supplied) (mL g-1), Y2P/S (mL g-1) (Utilized): (H2 mL/g glucose utilized), [Biomass] (g L-1). Biomass production g per L culture, YP/X (ml/g/L): (H2 ml per g Biomass per L), YX/S: (Biomass production per g glucose supplied), YH2/s: (conversion of H2 mL to H2 (g) per g glucose utilized) Glucose: 0.5 g L-1, inoculum size 10% (v/v), I pH. 7.0, Temp. 30°C | |
The data shown in Table 2 and 4 were used to Calculate different yields like YP/X (ml/g/L): (H2 mL per g Biomass per L), YX/S: (Biomass production per g glucose supplied) and YH2/s (conversion of H2 (ml) to H2 (g) per g glucose utilized). These results showed that fermentation medium without salts addition resulted to enhanced the bacterial growth and that would enhance the bacterial productivity of hydrogen. Suggested that the enhancement of hydrogen production due to the increased in bacterial biomass.
Mechanisms of sodium chloride inhibition on h2 production: Das and Veziroglu (2001) they mentioned that many ions can be inhibitory or toxic to H2 producing microorganisms, depending on their concentration. The results obtained in Table 1 and 2 showed that both H2 productivity and yield influenced by addition of NaCl to fermentation medium. Zheng et al. (2005) reasoned that to the ionic strength of the medium. They reported that a trace level of many ions is required for activation of function of many enzymes and co-enzymes. Excessive amounts, however, can lead to inhibition or toxicity. This is mostly due to chemical binding of some metals to the enzymes, resulting in the disruption of enzyme structure and activities. However, the inhibitory effect of NaCl on hydrogen production should not be related to the disruption of structure and activities of enzymes responsible for H2 production, as NaCl dose not bind to the enzyme. With high concentration of NaCl, high osmolarity environments trigger rapid fluxes of cell water, thus causing a reduction in turgor and dehydration of the cytoplasm (Ahring and Westermann, 1983).
In earlier studies about the effect of NaCl on methanogenic microorganisms, Zheng et al. (2005) stated that high osmolarity has been found to be responsible for the inhibition of NaCl on bacterial bioactivities. This was also likely to be the mechanism for NaCl inhibition of hydrogen production in the present study. Lin and Lay (2005) reported that magnesium, sodium, zinc and iron were important trace metals to enhanced hydrogen production by mixed culture with magnesium being the most significant. To find out the effect of sodium ions on the enzymes activity, Maris et al. (1984) study the levels of enzymes involved in the formation of acetate and butyrate in C. acetobutylicum. They found that in the pathway to form acetate, this bacterium produces two enzymes which are phosphate acetyltransferase and acetate kinase, they found that the activity of phosphate acetyltransferase inhibited by monovalent cation like Na+. It seems that the medium components was totally responsible of the inhibition of hydrogen production and with relation to other study weather the addition was to activate or to inhibit depend on the microorganism (s) used. Finally, NaCl is one of yeast extract (Nitrogen source) composition so, it might be that has another inhibitory effect and reduce the quantity of produced gases. In present study the results show to the first time that NaCl with any concentration in fermentation medium have negative effect on hydrogen production by C. acetobutylicum NCIMB13357.
Inhibition by sodium acetate: The results shown in Table 3 and 4 demonstrated that hydrogen production by C. acetobutylicum NCIMB13357, was depending on sodium acetate concentration and as Sodium Acetate concentration increased the hydrogen production yield and that connected to bacterium productivity of hydrogen were decreased. As mentioned above for sodium chloride, Niel et al. (2003) reasoned that inhibition of decreased hydrogen production to increase in concentration of the acetate in the fermentation medium and that affect the bacterium metabolic pathway. The effect of sodium acetate concentration it was also shown on the growth of bacteria. Growth was checked by final biomass concentration (bacterial dry weight). It appeared that concentration of inhibitor affect the bacterial growth and as concentration increased, the biomass concentration was decreased. Niel et al. (2003) suggested that due to cell lysis.
It was reported by Chia et al. (1982) the summary of earlier reports of the effect of sodium acetate salt on yeast growth. They reported that elevated levels of inorganic electrolytes in the liquid growth medium have been found to influence several parameters of yeast activity. (i) Cell growth and multiplication: (a) the number of viable yeast cells per unit volume of liquid growth medium decreases as salt content increases, (b) the biomass of the culture (i.e., the total weight of yeast cells per unit volume of liquid growth medium) decreases as salt content increases and (c) the length of the lag phase (i.e., the incubation period between inoculation of the culture and detectable initiation of cell growth) lengthens as salt concentration increases. (ii) Utilization of the primary carbon and energy source is reduced. (iii) Change in concentration of metabolic products: (a) there is a decrease in the production of ethanol as salt content increases and (b) there is an increase in the concentration of other fermentation products (such as glycerol, acetaldehyde, etc.) as salt content increases. Another study regarding the effect of sodium acetate on the enzyme level was conducted by Biggins and Dilworth (1968) they reported that an increase in the concentration of acetate within the cell to partially inhibit acetate kinase activity, resulting in elevated concentrations of acetyl phosphate in C. pasteurianum. They found also that acetyl phosphate acted as a product inhibitor of pyruvate phosphoroclastic activity and apparently resulted in a slowdown of general metabolism. Regarding the effect of sodium acetate on bacterial metabolism, Bechtle (1969) reported that sodium acetate was shown to have a potentiating effect on some proteases in hydrolysis of milk proteins at acid pH values and that effect was minimal in neutral pH media and there was an inhibition of the protease activity at alkaline pH values under some conditions. Furthermore, Lawrence and Sanderson (1969) reported that sodium acetate in the Cheesman assay medium was inhibitory to some of the microbial enzymes tested. This, as well as observations reported here, would indicate that acetate ions may have an inhibitory action on an enzyme system.
Inhibition of growth by acetate and other weak organic acids is quite common, but the level of tolerance varies tremendously among species. The undissociated form of these compounds can function as uncoupling agents (Booth, 1985). However, Niel et al. (2003) reported that, the ionic strength of sodium acetate was responsible for the inhibition of the hydrogen production by C. saccharolyticus and Donnison et al. (1989) observed that lysis in exponentially growing cultures of this organism after addition of 20 mM sodium acetate. Similar phenomena have already been described for, C. thermoaceticum (Wang and Wang, 1984) and Bacillus subtilis (Joliffe et al., 1981). Joliffe et al. (1981) reported that C. thermoaceticum release of autolysins upon addition of high salt concentrations. Another study by Kemper et al. (1993), they reported that inactive autolysins are distributed in the relatively acidic cell wall of gram-positive bacteria during exponential growth. Suggested that it is thought that autolysins become activated once the pH of the cell wall is neutralized. Results obtained in this study imply a similar scenario in C. saccharolyticus, which possesses also a gram-positive-type cell wall. Present results shows to the first time that sodium with any concentration in fermentation medium have negative effect on hydrogen production by bacterium we used in this study.
Finally the final H2 yield obtained by using the new medium was 340 mL g-1 glucose supplied (2.72 mol H2/mol glucose supplied): (According to Wooshin et al. (2005): each 125 mL of H2 ∼ 1 mole H2) and that was higher than reported value in the literature for mesophilic species of clostridia and using glucose as substrate. The entire reported H2 yield reported in the literature shown in Table 5. Through the revision of the components of the medium they were used, it was shown that some they used NaCl and different nitrogen source with different concentration and that what it discovered to inhibit the H2 production by C. acetobutylicum NCIMB13357 in this study. For above reasons it was clear that the medium composition had remarkable effect on hydrogen production by Clostridium species.
| Table 5: | Some of reported value of H2 by Clostridium species through the literature |
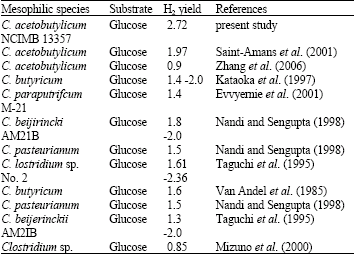 | |
| H2 yield (molH2/mol glucose supplied) | |
CONCLUSIONS
This study demonstrated fermentative hydrogen production by C. acetobutylicum NCIMB 13357 in a batch bioreactor. The maximum hydrogen yield was 391 mL g-1 glucose utilized. It found that both NaCl and sodium acetate were inhibitor to hydrogen production as well as bacterial growth of bacterium used in this study.
ACKNOWLEDGMENT
The author would like the thank Universiti Kebangsaan Malaysia for financial assistance under grant No UKM-OUP-BTK-14/2007.
REFERENCES
- Ahring, B.K. and P. Westermann, 1983. Toxicity of heavy metals to thermophilic anaerobic bacteria digestion. Applied Microbial. Biotechnol., 17: 365-370.
CrossRefDirect Link - Bechtle, R.M., 1969. Effect of sodium acetate on protease activity. J. Dairy Sci., 52: 1733-1737.
CrossRef - Biggins, D.R. and M.J. Dilworth, 1968. Control of pyruvate phosphoroclastic activity in extracts of Clostridium pasteurianum by ADP and acetyl phosphate. Biochim. Biophys. Acta, 156: 285-296.
PubMedDirect Link - Booth, I.R., 1985. Regulation of cytoplasmic pH in bacteria. Microbiol. Rev., 49: 359-378.
Direct Link - Chia-Jenn, W., D. Robert, D. Tanner, W. George and M. Malaney, 1982. Effect of sodium chloride on bakers yeast growing in gelatin. Applied Environ. Microbiol., 43: 757-763.
Direct Link - Das, D. and T.N. Veziroglu, 2001. Hydrogen production by biological processes: A survey of literature. Int. J. Hydrogen Energy, 26: 13-28.
CrossRef - Joliffe, L.K., R.J. Doyle and U.N. Streips, 1981. The energized membrane and cellular autolysis in Bacillus subtilis. Cell, 25: 753-763.
CrossRef - Kataoka, N., A. Miya and K. Kiriyama, 1997. Studies on hydrogen production by continuous culture system of hydrogen producing anaerobic bacteria. Water Sci. Technol., 36: 41-47.
Direct Link - Kemper, M.A., M.M. Urrutia, T.J. Beveridge, A.L. Koch and R.J. Doyle, 1993. Proton motive force regulate cell wall-associated enzymes of Bacillus subtilis . J. Bacteriol., 175: 5690-5696.
PubMed - Lawrence, R.C. and W.B. Sanderson, 1969. A micro-method for the quantitative estimation of rennets and other proteolytic enzymes. J. Dairy Res., 36: 21-21.
Direct Link - Lee, Y., T. Miyahara and T. Noike, 2001. Effect of iron concentration on hydrogen fermentation. Bioresour Technol., 80: 227-231.
CrossRef - Lin, C.Y. and C.H. Lay, 2005. A nutrient formulation for fermentative hydrogen production using anaerobic sewage sludge microflora. Int. J. Hydrogen Energy, 30: 285-292.
CrossRef - Maris, G., N. Hartmanis and G. Sten, 1984. Intermediary Metabolism in Clostridium acetobutylicum: Levels of enzymes involved in the formation of acetate and butyrate. Appl. Environ. Microbiol., 47: 1277-1283.
PubMedDirect Link - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Mizuno, O., R. Dinsdale, F.R. Hawkes, D.L. Hawkes and T. Noike, 2000. Enhancement of hydrogen production from glucose by nitrogen gas sparging. Bioresour. Technol., 73: 59-65.
CrossRefDirect Link - Morimoto, M., M. Atsuko, A.A.Y. Atif, M.A. Ngan and A. Fakhrul-Razi et al., 2004. Biological production of hydrogen from glucose by natural anaerobic microflora. Int. J. Hydrogen Energy, 29: 709-713.
CrossRefDirect Link - Nandi, R. and S. Sengupta, 1998. Microbial production of hydrogen: An overview. Crit. Rev. Microbiol., 24: 61-84.
CrossRefPubMedDirect Link - Van Andel, J.G., G.R. Zoutberg, P.M. Crabbendam and A.M. Breure, 1985. Glucose fermentation by Clostridium butyricum grown under a self generated gas atmosphere in chemostat culture. Applied Microbiol. Biotechnol., 23: 21-26.
CrossRef - Wang, G. and D. Wang, 1984. Elucidation of growth inhibition and acetic acid production by Clostridium thermoaceticum . Applied Environ. Microbiol., 47: 294-298.
PubMedDirect Link - Xiaolong, H., Z. Minghua, Y. Hanqing, S. Qinqin and Lei Lecheng, 2006. Effect of sodium ion concentration on hydrogen production from sucrose by anaerobic hydrogen-producing granular sludge. Chinese J. Chem. Eng., 14: 511-517.
Direct Link - Zhang, H.B., A. Mary and E.L. Bruce, 2006. Biological hydrogen production by Clostridium acetobutylicum in an unsaturated flow reactor. Water Res., 40: 728-734.
CrossRef