
H.R. Asghari
Faculty of Agriculture, Shahrood University of Technology, P.O. Box 3619995161-316, Shahrood, Iran
M.R. Amerian
Faculty of Agriculture, Shahrood University of Technology, P.O. Box 3619995161-316, Shahrood, Iran
H. Gorbani
Faculty of Agriculture, Shahrood University of Technology, P.O. Box 3619995161-316, Shahrood, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 15 | Page No.: 1909-1915







ABSTRACT
In order to determine the effects of soil salinity on AM fungi colonization in halophytes, plants of semi-arid region of North-Eastern Iran were examined for their colonization in soils with different salinity levels. Roots of several halophytes were colonized and showed typical structure of AM fungi with different levels of colonization. Haloxylon aphyllum, Kochia stellaris, Halocnemum strobilaceum, Seidlitzia rosmarinus and Salsola sp. of the Chenopodiaceae and Zygophyllum eurypterum and Peganum harmala of the Zygophyllaceae were found to be colonized by AM fungi. In several species the mycorrhizal status is reported for the first time. The results of this study revealed that AM colonization in halophytes in soil with high salinity level (16 dS m-1), but colonization was inhibited by very high salinity (45 dS m-1). The AM fungi colonization was absent in halophytes in very high soil salinity conditions may was due to inability of AM fungi to survive such salinity conditions, which may limit the beneficial effects of AM fungi in halophytes.
PDF Abstract XML References Citation
How to cite this article
H.R. Asghari, M.R. Amerian and H. Gorbani, 2008. Soil Salinity Affects Arbuscular Mycorrhizal Colonization of Halophytes. Pakistan Journal of Biological Sciences, 11: 1909-1915.
DOI: 10.3923/pjbs.2008.1909.1915
URL: https://scialert.net/abstract/?doi=pjbs.2008.1909.1915
DOI: 10.3923/pjbs.2008.1909.1915
URL: https://scialert.net/abstract/?doi=pjbs.2008.1909.1915
INTRODUCTION
Mycorrhizal symbiosis is an association between fungi and plant roots. In this association the fungi provide a fundamental link between soil and plant roots which could increase transport and nutrient uptake in plants through the extension of fungal hyphae in soil. Mycorrhizal symbiosis is found in average on 80-90% of land plants in different soil conditions (Smith and Read, 1997). Absence of mycorrhizal fungi was reported in halophytes species mostly belonging to the Chenopodiaceae family in early studies (Hirrel et al., 1978).
Arbuscular mycorrhizal fungi occur in many stressful environments. Relatively large populations of AM fungi spores have been found in saline conditions (Aliasgharzadeh et al., 2001; Sengupta and Chaudhuri, 1990; Wang et al., 2004). The results of glasshouse studies have shown that AM fungi can increase plant salinity tolerance and yield in saline conditions (Al-Karaki, 2006; Asghari et al., 2005; Hirrel and Gerdemann, 1980; Ojala et al., 1983; Plenchette and Duponnois, 2005; Tian et al., 2004), but salinity may have negative effects on AM fungi growth and hyphal extension (Juniper and Abbott, 1993; Peat and Fitter, 1993).
The distribution of AM fungi in saline soils has been investigated in many studies (Aliasgharzadeh et al., 2001; Hildebrandt et al., 2001; Garcia and Mendoza, 2008; Landwehr et al., 2002; Sengupta and Chaudhuri, 1990; Wang et al., 2004). Aliasgharzadeh et al. (2001) reported that increasing soil salinity decreased the percentage of AM colonization in glycophytes. Garcia and Mendoza (2008) found high level of AM colonization in saline soils of a temperate grassland, which water content, salinity and sodicity in soil were positively associated with AM root colonization and arbuscule colonization in Lotus tenuis, but negatively so in the grasses The effects of soil salinity on spore germination of AM fungi and therefore hyphal production is one of the most important detrimental effects of salinity on mycorrhizal colonization (Juniper and Abbott, 2006).
Different levels of AM colonization in halophytes have been reported in many field studies in different locations, seasons and soil salinity levels, but AM colonization in halophytes along a salinity gradient has received less attention. The objectives of this study were to evaluate AM colonization of halophytes and also to investigate the effects of increased soil salinity along a transect on AM fungi colonization in halophytes of semi-arid region of North-Eastern Iran.
MATERIALS AND METHODS
Study site: This study was conducted in 2005 at Turan Biosphere Reserve (TBR), situated in north east of Iran (Fig. 1). The reserve includes 1.8 million hectares of flat, semi-arid desert plains which was set aside for conservation and research by the United Nations Development Program (UNDP) in 1972. The climate is semi-arid, with dry summers and cold winters. The 20-year mean annual rainfall is 110-170 mm. Rainfall varies across the TBR, decreasing from north to South.
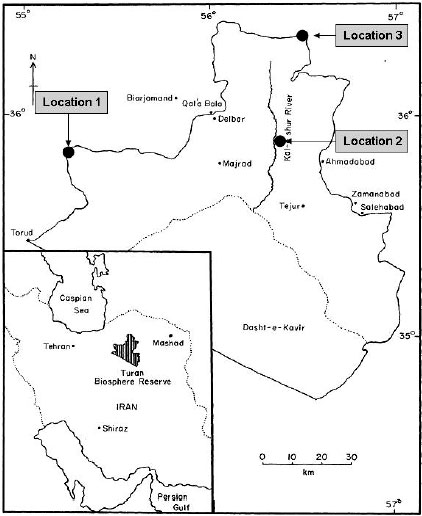 | |
| Fig. 1: | Turan biosphere reserve of North-Eastern Iran, showing the three survey locations |
Collection of plant specimens and soil: Two different sampling stages were carried out. At the first sampling event, three different locations in north, east and center of TBR (Fig. 1) were selected in June 2005 to study root AM colonization of different halophytes. The second sampling event was carried out along a 40 km north-south transect extending from a low soil salinity zone to a high soil salinity zone, in location 1 (Fig. 1) in April 2006 to study the effects of soil salinity on AM colonization in halophytes. At each location, soil and root samples were collected from 0-30 cm of three individual plants. Samples were taken from halophytes that had no immediately adjacent plants within approximately 1 m, to avoid contamination by roots of other plants. AM colonization is the pre-requisite for the interaction between AM fungi and plants and is therefore a better measure for the effect of AM on plants than spore counts. Therefore the percent of AM colonization of roots was estimated in this study and the density of spores in soil was not measured. AM colonization was assessed after clearing washed roots with 10% KOH for 3 days at room temperature and staining with trypan blue (Phillips and Hayman, 1970). Darkly pigmented roots were cleared with alkaline hydrogen peroxide (0.5% NH4OH and 0.5% H2O2 v/v in water) (Brundrett et al., 1996). After staining, AM colonization was determined in about 200-250 root segments by the gridline intersect method (Giovannetti and Mosse, 1980) .
Air-dry soil was crushed and sieved through a 2 mm mesh for particle-size analysis using the hydrometer method. The Total Neutralizing Value (TNV) as calcium carbonate and magnesium carbonate was measured by back titration procedure. Soil pH and salinity of water extract of samples were determined by pH meter and electrical conductivity meter, respectively.
RESULTS
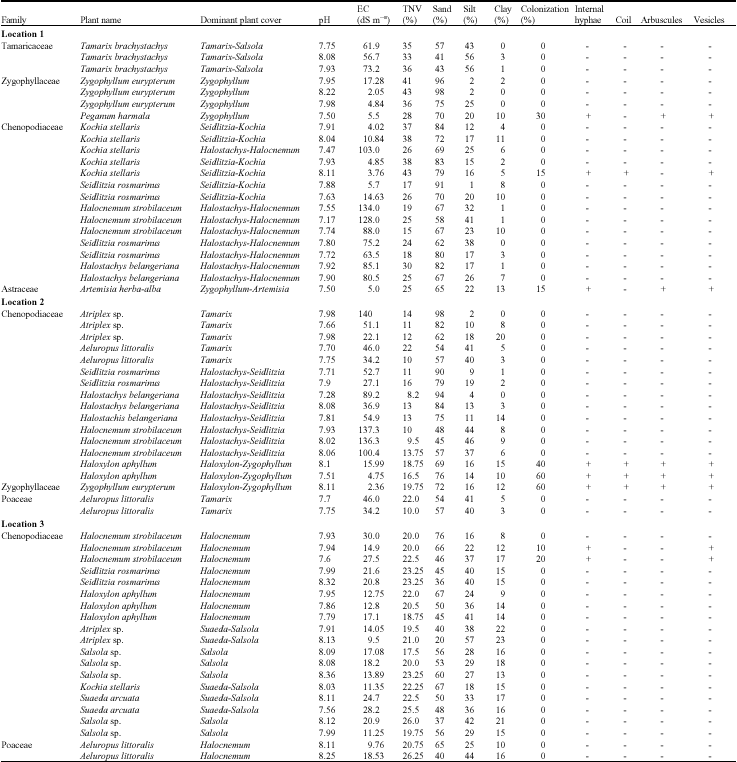
AM colonization in different locations: Of the plant species surveyed in first sampling event, 6 of 12 halophyte species (50%) showed some evidence of AM fungi colonization in at least one of the locations (Table 1). Members of Chenopodiaceae (Haloxylon aphyllum, Kochia stellaris and Halocnemum strobilaceum), Zygophyllaceae (Zygophyllum eurypterum and Peganum harmala) and Astraceae (Artemisia herba-alba) were all colonized. Except of Poaceae, AM fungi structures were found in all investigated plant families. The lowest percentages of AM colonization (10%) were observed in Halocnemum strobilaceum and the highest (60%) in Haloxylon aphyllum and Zygophyllum eurypterum. AM colonization in roots of halophytes was found in soil salinities range between 2.3-27.5 dS m-1. The highest AM colonization occurred at the low salinity levels. The same plant species had different levels of AM colonization in different locations at nearly the same salinity levels (see Haloxylon aphyllum in location 2 and 3 in Table 1). At the same salinity level and location different plant species were not equally colonized. For example at nearly the same salinity levels Peganum harmala was colonized 30% and no AM colonization were found in Zygophyllum eurypterum in location 1 (Table 1). The roots of halophytes in high salinities contained internal hyphae and vesicles but no other mycorrhizal structures.
| Table 1: | Arbuscular mycorrhizal fungi colonization of the roots of halophytes collected from Turan biosphere reserve of North-Eastern Iran |
 | |
| Table 2: | Arbuscular mycorrhizal colonization of the roots of halophytes collected along a north-south transect in location 2 from turan biosphere reserve of North-Eastern Iran |
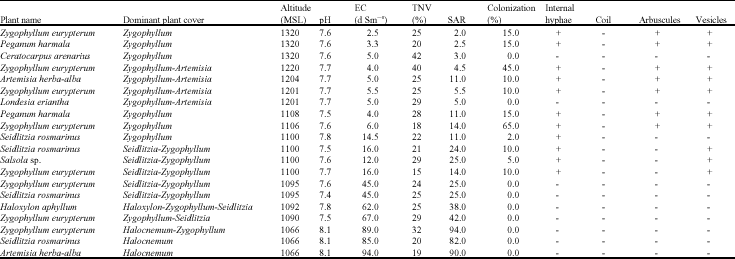 | |
Effects of soil salinity on AM fungi colonization along a north-south transect: Altitude decreased from north to south, soil salinity, pH and Sodium Absorption Ratio (SAR) increased with decreasing altitude, but no significant changes were found in soil total neutralizing value (TNV) (Table 2). AM colonization was observed only between 2.5 to 16 dS m-1 and no AM fungi structure were found in very high salinity levels (45-94 dS m-1) in all plant species investigated in this sampling. The percentage of AM colonization ranged from 2 to 65 in different plant species.
Zygophyllum eurypterum as a dominant plant along the transect, growing in different levels of soil salinity (2.5-89 dS m-1), showed different levels of AM fungi colonization. The results showed that all mycorrhizal structures were found in Zygophyllum eurypterum roots at soil salinity levels up to 6 dS m-1, but no arbuscules were found in soil salinities between 6 and 16 dS m-1 whereas no mycorrhizal structure were observed at 45 dS m-1. The same trend towards AM absence in increased salinity level was found in Artemisia herba-alba and Seidlitzia rosmarinus (Table 2).
DISCUSSION
Occurrence of AM colonization of some halophytes has been reported by researchers in field and glasshouse conditions (Aldon, 1975; Hildebrandt et al., 2001; Sengupta and Chaudhuri, 1990; Wang et al., 2004). The results of this study showed different levels of AM colonization with different halophytes host plants (mostly from Chenopodiaceae family) which is in agreement with previous field studies. To our knowledge, the occurrence of AM fungi colonization in roots of Zygophyllum eurypterum and Kochia stellaris is reported for the first time in this study (Fig. 2).
The results of the first sampling showed that plant species from different family had different levels of AM fungi colonization. Different plant species growing at the same salinity level were colonized differently, which suggests that AM colonization is, at least partly, regulated by the plant species. Also finding different levels of AM colonization in the same plants species at the same salinity levels in different locations indicate that other factors such as soil moisture, soil layer and soil chemical and physical properties than salinity and plant species control AM colonization (McMillen et al., 1998; Nadian et al., 1998; Wang et al., 2004).
Overall the results of the first and second sampling stages showed that higher percentage of AM colonization occur at lower soil salinity levels and very high soil salinity was linked to the absence of AM fungi colonization in roots of halophytes. Zygophyllum eurypterum grew at different salinity levels (2.5 to 89 dS m-1), but AM colonization did not occur at very high salinity levels (45 to 89 dS m-1). This result showed the effects of edaphic properties to control AM fungi colonization in this halophytic plant. The same results were reported by other researchers in some other species and lower salinities.
Salinity could reduce AM colonization by directly reducing hyphal growth and/or decreasing plant growth (less carbohydrate). A recent report indicates that the most important effect of salinity on AM fungi is related to its detrimental effect on spore germination and hyphal production (Juniper and Abbott, 2006). On the other hand, in AM associations the fungus is completely depend on plant growth and carbohydrate nutrition production in host plant, therefore every factor that effects the carbohydrate production and its translocation to the roots could effect the amount of mycorrhizal colonization (Thomson et al., 1990). Since salinity reduces plant growth and decreases carbohydrates concentration in host plant (Greenway and Munns, 1980), it could reduce mycorrhizal colonization. Poor growth of Zygophyllum eurypterum by increased soil salinity in location 1 has been observed, suggesting that the plant could not supply the AM fungi with enough carbohydrate.
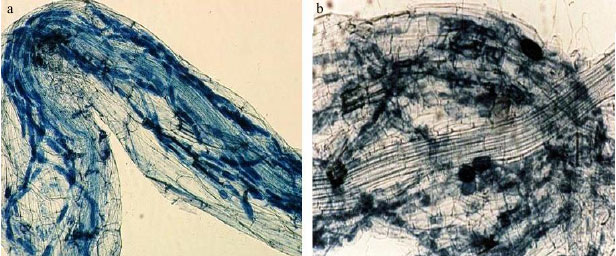 | |
| Fig. 2: | Roots of halophytes with arbuscular fungi mycorrhizal structures, (a) Zygophyllum eurypterum and (b) Kochia stellaris |
In previous studies around the world, AM fungi colonization in roots of halophytes was found in different levels of soil salinity up to 185 dS m-1. Different structures of AM colonization were found in some halophytes such as Atriplex canescences (Barrow et al., 1997), Tamarix chinesis and Aeluropus littoralis (Wang et al., 2004) and Suaeda maritime (Sengupta and Chaudhuri, 1990), but in this study no AM fungal structures were found in these genera of halophytes, even at relatively low salinity levels. However it is difficult to determine factors which actually control AM fungi colonization. Lack of root exudates (Nagahashi and Douds, 1999) or mycorrhizal helper microorganisms (Caroline and Bagyaraj, 1995) in the rhizosphere, presence of toxic compounds of root exudates (Peterson and Bradbury, 1995; Vierheilig et al., 1995) or intrinsic barrier of the root cortex or epidermis etc. Which have been reported previously for absence of AM colonization in plants in non saline conditions. Furthermore absence of salt tolerant AM fungi species could be the case in investigated soils with high salinity levels. The AM fungi which are isolated from saline soils which may have an ability to improve the survival of host plants (Copeman et al., 1996).
Moreover, different methods of root staining and times of investigation in a plant may cause different results. In this study some of halophytes species such as Haloxylon aphyllum, Halostachys belangeriana and Seidlitzia rosmarinus had dark roots, which did not clear completely making AM colonization difficult to observe. Different root clearing and staining methods is suggested in the future works.
The effects of AM fungi on plant salinity tolerance of a range of glycophytes have been shown previously. Furthermore AM fungi increased growth of halophytes in saline conditions in some glasshouse studies (Aldon, 1975; Asghari et al., 2005; Hirrel and Gerdemann, 1980; Plenchette and Duponnois, 2005). It is speculated that AM fungi improve plant growth in both glycophytes and halophytes in saline conditions. The results of this field study showed that high soil salinity lead to the absence of AM colonisation, which indicate halophytes can grow in higher soil salinity levels than AM fungi. This finding can show the limitation of AM fungi use in halophytes in very saline soils. These results may important in application of AM fungi in halophytes in revegetation of saline soils.
ACKNOWLEDGMENTS
Support for this research was provided by Shahrood University of Technology, Iranian Ministry of Science, Research and Technology. The author`s thanks Mr. M.A. Hoseinpour, Mr. G.R. Shakeri, Ms. A. Hassani and Ms. S. Ahmadi for their excellent technical assistance and authors thanks members of faculty of Agriculture of Shahrood University of Technology for their helpful discussions.
REFERENCES
- Aliasgharzadeh, N., S.N. Rastin, H. Towfighi and A. Alizadeh, 2001. Occurrence of arbuscular mycorrhizal fungi in saline soils of the Tabriz Plain of Iran in relation to some physical and chemical properties of soil. Mycorrhiza, 11: 119-122.
CrossRefDirect Link - Al-Karaki, G.N., 2006. Nursery inoculation of tomato with arbuscular mycorrhizal fungi and subsequent performance under irrigation with saline water. Sci. Hortic., 109: 1-7.
CrossRefDirect Link - Asghari, H.R., P. Marschner, S.E. Smith and F.A. Smith, 2005. Growth response of Atriplex nummularia to inoculation with arbuscular mycorrhizal fungi at different salinity levels. Plant Soil, 273: 245-256.
CrossRefDirect Link - Barrow, J.R., K.M. Havstad and B.D. McCaslin, 1997. Fungal root endophytes in fourwing saltbush, Atriplex canescens, on arid rangelands of Southwestern USA. Arid Soil Res. Rehabil., 11: 177-185.
Direct Link - Brundrett, M., N. Bougher, B. Dell, T. Grove and N. Malajczuk, 1996. Working with Mycorrhizas in Forestry and Agriculture. Australian Centre for International Agricultural Research, Canberra, Australia, ISBN: 1862301815, Pages: 374.
Direct Link - Copeman, R.H., C.A. Martin and J.C. Stutz, 1996. Tomato growth in response to salinity and mycorrhizal fungi from saline or non-saline soils. Hortscience, 31: 341-344.
Direct Link - Garcia, I.V. and R.E. Mendoza, 2008. Relationships among soil properties, plant nutrition and arbuscular mycorrhizal fungi�`plant symbioses in a temperate grassland along hydrologic, saline and sodic gradients. FEMS Microbiol. Ecol., 63: 359-371.
CrossRef - Giovannetti, M. and B. Mosse, 1980. An evaluation of techniques to measure vesicular-arbuscular infection in roots. New Phytol., 84: 489-500.
CrossRefDirect Link - Greenway, H. and R. Munns, 1980. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol., 31: 149-190.
CrossRefDirect Link - Hildebrandt, U., K. Janetta, F. Ouziad, B. Renne, K. Nawrath and H. Bothe, 2001. Arbuscular mycorrhizal colonization of halophytes in Central European salt marshes. Mycorrhiza, 10: 175-183.
CrossRefDirect Link - Hirrel, M.C. and J.W. Gerdemann, 1980. Improved growth of onion and bell pepper in saline soils by two vesicular-arbuscular mycorrhizal fungi. Soil Sci. Am. J., 44: 654-658.
Direct Link - Hirrel, M.C., H. Mehravaran and J.W. Gerdemann, 1978. Vesicular-arbuscular mycorrhiza in the Chenopodiaceae and Cruciferae: do they occur?. Can. J. Bot., 56: 2813-2817.
CrossRefDirect Link - Juniper, S. and L. Abbott, 1993. Vesicular-arbuscular mycorrhizas and soil salinity. Mycorrhiza, 4: 45-57.
CrossRefDirect Link - Juniper, S. and L.K. Abbott, 2006. Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi. Mycorrhiza, 16: 371-379.
CrossRefDirect Link - Landwehr, M., U. Hildebrandt, P. Wilde, K. Nawrath, T. Toth, B. Biro and H. Bothe, 2002. The arbuscular mycorrhizal fungus Glomus geosporum in European saline, sodic and gypsum soils. Mycorrhiza, 12: 199-211.
CrossRefPubMedDirect Link - McMillen, B.G., S. Juniper and L.K. Abbott, 1998. Inhibition of hyphal growth of a vesicular-arbuscular mycorrhizal fungus in soil containing sodium chloride limits the spread of infection from spores. Soil Biol. Biochem., 30: 1639-1646.
Direct Link - Nadian, H., S.E. Smith, A.M. Alston, R.S. Murray and B.D. Siebert, 1998. Effects of soil compaction on phosphorus uptake and growth of Trifolium subterraneum colonized by four species of vesicular-arbuscular mycorrhizal fungi. New Phytol., 140: 155-165.
CrossRefDirect Link - Nagahashi, G. and D.D. Douds, 1999. Rapid and sensitive bioassay to study signals between root exudates and arbuscular mycorrhizal fungi. Biotechnol. Tech., 13: 893-897.
CrossRefDirect Link - Ojala, J.C., W.M. Jarrell, J.A. Menge and E.L.V. Johnson, 1983. Influence of mycorrhizal fungi on the mineral nutrition and yield of onion in saline soil. Agron. J., 75: 255-259.
Direct Link - Peat, H.J. and A.H. Fitter, 1993. The distribution of arbuscular mycorrhizas in the British flora. New Phytol., 125: 845-854.
CrossRefDirect Link - Peterson, R.L. and S.M. Bradbury, 1995. Use of Plant Mutants, Intraspecific Variants and Non-Hosts in Studying Mycorrhiza Formation and Function. In: Mycorrhiza, Structure, Function, Molecular Biology and Biotechnology. Hock, B. (Ed.). Springer Verlag, Berlin Heidelberg, ISBN: 978-3540639817 pp: 157-180.
- Phillips, J.M. and D.S. Hayman, 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc., 55: 158-161.
CrossRefDirect Link - Plenchette, C. and R. Duponnois, 2005. Growth response of the saltbush Atriplex nummularia L. to inoculation with the arbuscular mycorrhizal fungus Glomus intraradices. J. Arid Environ., 61: 535-540.
CrossRefDirect Link - Sengupta, A. and S. Chaudhuri, 1990. Vesicular arbuscular mycorrhiza (VAM) in pioneer salt marsh plants of the Ganges river delta in West Bengal (India). Plant Soil, 122: 111-113.
CrossRefDirect Link - Smith, S.E. and D.J. Read, 1997. Mycorrhizal Symbiosis. 2nd Edn., Academic Press, London, UK., ISBN-13: 978-0-12-652840-4, Pages: 605.
Direct Link - Tian, C.Y., G. Feng, X.L. Li and F.S. Zhang, 2004. Different effects of arbuscular mycorrhizal fungal isolates from saline or non-saline soil on salinity tolerance of plants. Applied Soil Ecol., 26: 143-148.
CrossRefDirect Link - Vierheilig, H., M. Alt, J. Lange, M. Gut-Rella, A. Wiemken and T. Boller, 1995. Colonization of transgenic tobacco constitutively expressing pathogenesis-related proteins by the vesicular-arbuscular mycorrhizal fungus Glomus mosseae. Applied Environ. Microbiol., 61: 3031-3034.
Direct Link - Wang, F.Y., R.J. Liu, X.G. Lin and J.M. Zhou, 2004. Arbuscular mycorrhizal status of wild plants in saline-alkaline soils of the Yellow River Delta. Mycorrhiza, 14: 133-137.
CrossRefDirect Link - Thomson, B.D., A.D. Robson and L.K. Abbott, 1990. Mycorrhizas formed by Gigaspora calospora and Glomus fasciculatum on subterranean clover in relation to soluble carbohydrate concentrations in roots. New Phytol., 114: 217-225.
CrossRef