
N. Agh
Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Ghent, Belgium
2Artemia and Aquatic Animals Research Institute, Urmia University, Urmia, Iran
G. Van Stappen
Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Ghent, Belgium
P. Bossier
Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Ghent, Belgium
H. Sepehri
Department of Biology, Faculty of Science, Tehran University, Tehran, Iran
V. Lotfi
Artemia and Aquatic Animals Research Institute, Urmia University, Urmia, Iran
S.M. Razavi Rouhani
Faculty of Veterinary Sciences, Urmia University, Urmia, Iran
P. Sorgeloos
Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Ghent, Belgium
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 2 | Page No.: 164-172







ABSTRACT
This study deals with effects of different salinities on the survival, growth, reproductive and lifespan characteristics of three Artemia populations from Urmia Lake and small lagoons at the vicinity of the lake under laboratory conditions. Experimental salinities ranged from 75 to 175 g L-1. Salinity was proved to have significant impact on the majority of the characters studied in this survey. Growth and survival in bisexual A. urmiana and parthenogenetic Artemia from Lake Urmia were significantly higher with respect to the parthenogenetic Artemia from lagoons at most of the salinities tested. Reproductive characteristics such as total number of broods, total offspring number of offspring in each brood and number of offspring at each day of reproductive period reduced with increasing salinity. Moreover higher salinity prolonged the pre-reproductive period but shortened the total reproductive period. Higher salinities also affected the percentage of encystment and post-reproductive period, showing significantly higher values in parthenogenetic populations in comparison to bisexual A. urmiana.
PDF Abstract XML References Citation
How to cite this article
N. Agh, G. Van Stappen, P. Bossier, H. Sepehri, V. Lotfi, S.M. Razavi Rouhani and P. Sorgeloos, 2008. Effects of Salinity on Survival, Growth, Reproductive and Life Span Characteristics of Artemia Populations from Urmia Lake and Neighboring Lagoons. Pakistan Journal of Biological Sciences, 11: 164-172.
DOI: 10.3923/pjbs.2008.164.172
URL: https://scialert.net/abstract/?doi=pjbs.2008.164.172
DOI: 10.3923/pjbs.2008.164.172
URL: https://scialert.net/abstract/?doi=pjbs.2008.164.172
INTRODUCTION
The brine shrimp Artemia is a genus with a wide distribution on the five continents, inhabiting inland salt lakes, coastal lagoons and solar saltworks (Vanhaecke et al., 1987). It is among the unique organisms that can adapt to very diverse living conditions that involve salinities as low as 10 g L-1 (Abatzopoulos et al., 2006a) to as high as 340 g L-1 (Post and Youssef, 1977). It comprises a complex of sibling species and superspecies defined by a criterion of reproductive isolation (Browne and Bowen, 1991).
Lake Urmia with about 5000 km2 is one of the largest permanent water catchments in West Asia. Lake Urmia is a thalassohaline, sodium chloride lake (Loffler, 1961) with oligotrophic characteristics, located at an altitude of 1250 m above sea level (Cole and Brown, 1967; Azari Takami, 1993). Its surface area was reported to range from 4750 to 6100 km2 and the average and greatest depths were between 6 and 16 m, respectively (Azari Takami, 1993; Van Stappen et al., 2001). However, according to recent studies by Agh (2006) the surface area of the lake has reduced to less than 4000 km2 and the average and greatest depths are between 3 and 6 m, respectively, due to the prolonged drought since 1999 and the construction of a number of reservoirs/dams on major inflowing rivers. Lake Urmia water salinity that used to fluctuate from 140 to 220 g L-1 before 1999, has never dropped below 250 g L-1 since then and is in saturation state almost throughout the year (Agh, 2006).
A. urmiana was first reported in Lake Urmia by Gunther in 1899. Many other researchers confirmed presence of this bisexual species of Artemia in Lake Urmia (Clark and Bowen, 1976; Barigozzi, 1989; Pador, 1995; Sorgeloos, 1997; Van Stappen et al., 2001; Agh, 2002; Noori and Agh, 2002). Most recently Agh et al. (2007) confirmed that a parthenogenetic population of Artemia coexists with the bisexual A. urmiana in Lake Urmia.
A parthenogenetic population of Artemia was reported from small lagoons at the vicinity of the Lake Urmia by Agh and Noori (1997). These lagoons are scattered at the periphery of the lake in both West and East Azerbaijan. The size of the lagoons varies from a few square meters to maximum 10000 m2 surface area and their depth is always less than 0.7 m. Therefore these lagoons are considered as temporary small water catchments that are dried during early summer and filled up again during winter rains. Water salinity in the lagoons ranges from 10-20 g L-1 in early spring and gradually rises to saturation level within about 10 weeks. Parthenogenetic females were observed at high densities with rare males seen only at the ratio of one male to 100 females in these lagoons (Agh and Noori, 1997; Abatzopoulos et al., 2006a). Parthenogenetic Artemia in these lagoons grow to maturity at very low salinities (as low as 10 g L-1) and start reproduction at salinity of 15-20 g L-1 (Agh et al., 2007). The lagoons get saturated with brine within this period, but still a very high density of dark red coloured Artemia could be observed in these lagoons before they finally dry up.
There is considerable literature information on survival, growth, morphometry, reproductive and life span characteristics of many bisexual and parthenogenetic Artemia populations (Vanhaecke et al., 1984; Wear and Haslett, 1986; Browne et al., 1984, 1991; Browne and Wanigasekera, 2000; Triantaphyllidis et al., 1995, 1997a, b; Baxevanis et al., 2004; El-Bermawi et al., 2004; Abatzopoulos et al., 2003, 2006b). Most of these studies have contributed to the evaluation of genetic and environmental components of variance in sexual and or clonal Artemia. They have also enabled the comparison of life history characteristics and strategies between different populations (Browne et al., 2002; Abatzopoulos et al., 2003; Baxevanis and Abatzopoulos, 2004; Kappas et al., 2004). However, the effects of salinity on A. urmiana and parthenogenetic populations from Iran have been poorly examined or not studied at all.
In this study, we will survey the effect of salinity on growth, survival, reproductive and life span characteristics of three Artemia populations occurring in Urmia Lake region in order to understand their adaptation patterns to various salinities, thereby providing additional data on the characterization of Artemia populations from Iran.
MATERIALS AND METHODS
Culture procedure: This study was conducted at Artemia and Aquatic Animals Research Institute, Iran from June to December. 2004. Cysts of Artemia from the Lake Urmia and from the lagoons around the lake (Table 1) were hatched under standard conditions (Sorgeloos et al., 1986). The larvae hatched from the cyst samples of each biotope were siphoned into separate beakers and then transferred into separate 30 L aquaria, where they were cultured until adulthood. One hundred sexually matured bisexual couples (A. urmiana; URM) and equal number of adult parthenogenetic female Artemia (P-URM) were separated from the Urmia Lake sample on the basis of their different morphology and similarly 100 parthenogenetic females from the lagoon sample (LAG) were all transferred into separate 1.5 L cylindro-conical containers. The adult bisexual and parthenogenetic Artemia were allowed to reproduce inside the cones and the newly produced cysts and live nauplii were eliminated every alternate day until 2 weeks. This ensures that each population reproduces in
| Table 1: | List of the populations studied and abbreviations used |
 | |
its respective pattern of reproduction, sexually or asexually. This step was then followed by transferring six replicates of 200 actively moving nauplii from each population into 500 mL conical containers filled with 400 mL of filtered water with different salinities (50, 75, 100, 125, 150 and 175 g L-1). The animals were cultured at 27±1°C under constant aeration. The salinity in each cone was checked twice a day in order to maintain salinities according to the experimental set up. Artemia were fed unicellular algae Dunaliella tertiolecta and chemically treated yeast Lansy PZ® (INVE Technologies, Baasrode, Belgium) adopted from Coutteau et al. (1992).
Density of Artemia was adjusted to one animal/mL at the beginning of the experiment, but the density was gradually decreased to one animal per 3 mL on day 8 and per 4 mL on day 14 of growth.
Determination of survival and growth: Survival and total length were determined on days 8, 11, 14, 17, 20 and 23, according to Triantaphyllidis et al. (1995). For determining the growth 10 animals from each replicate were measured from the most anterior part of the head up to the last abdominal segment (telson) using a light microscope equipped with a phototube and micrometer. Drawings were later digitized using a digitizer connected to a computer.
Reproductive and life span characteristics: As animals attained maturity, 30 pairs of coupling bisexual A. urmiana and 30 adult parthenogenetic females from each population (from all salinities) were transferred into separate 50 mL falcon tubes in order to study their life cycle characteristics. Separate culture of individuals or couples continued as long as the female Artemia were alive. In the Falcon tubes containing bisexual A. urmiana, dead males were immediately replaced with actively swimming males during the experiment (Browne et al., 1988). The Falcon tubes were checked every day for the production of cysts or nauplii, which were counted and recorded separately. Finally, the reproductive characteristics (number of broods, total amount of offspring, brood size, offspring/day during the reproductive period, brood intervals and percentage of encystment) and the life span characteristics (pre-reproductive period, reproductive period, post-reproductive period and life span) were determined for each population according to Browne et al. (1984 ,1988). The results were statistically analyzed using SPSS (version 13) analysis of variance (ANOVA) (Sokal and Rohlf, 1981; Triantaphyllidis et al., 1995) and the averages were compared using Duncan`s test.
RESULTS
Survival: Records of the survival after 23 days of experiment indicate that in the majority of the cases survival declines in all populations when salinity increases. P-URM had significantly higher survival at salinities 75, 100 and 125 g L-1 compared to URM and LAG (Table 2, ANOVA, p<0.05). The bisexual URM had significantly higher survival only at 150 g L-1 compared to the asexual populations. No significant differences were observed in the survival of the three populations at 175 g L-1. Maximum survival of URM was observed at 100 g L-1, significantly higher compared to the values obtained at other salinities at intrapopulation level (Table 2, ANOVA, p<0.05), while minimum survival for this population was observed at 175 g L-1. No significant differences were observed in survival of URM at 75, 125 and 150 g L-1 salinities. The two asexual populations demonstrated significant differences in survival at all salinities.
Growth: The bisexual URM had the highest growth values in all salinities compared to the asexual populations (Table 3, ANOVA, p<0.05). The maximum total length of URM was significantly larger than this of the parthenogenetic strains (URM: 12.7±0.5 mm while for the asexual strains it was less than 9.7 mm). Total lengths of URM were almost similar at 75 and 100 g L-1 (Table 3) while this was significantly smaller at the three higher salinities (ANOVA, p<0.05).
No considerable differences were found in the total length of the parthenogenetic strains (P-URM and LAG) at 75 and 100 g L-1 (Table 3), but the same were significantly different at 125, 150 and 175 g L-1. (ANOVA, p<0.05). Maximum total length for P-URM (9.7 mm) was observed at 125 g L-1 while maximum growth for LAG (9.1 mm) was seen at 75 and 100 g L-1.
Reproductive and life span characteristics: At 75 g L-1, statistical analysis revealed that the bisexual strain (URM) was different from the parthenogenetic ones in six out of eleven measured characteristics (Table 4). However, URM had higher values only in the total number of nauplii produced, offspring per brood and post-reproductive period compared to those of parthenogenetic strains (P-URM and LAG). URM had significantly lower values
| Table 2: | Mean value±SD of survival percentage of three Artemia populations studied |
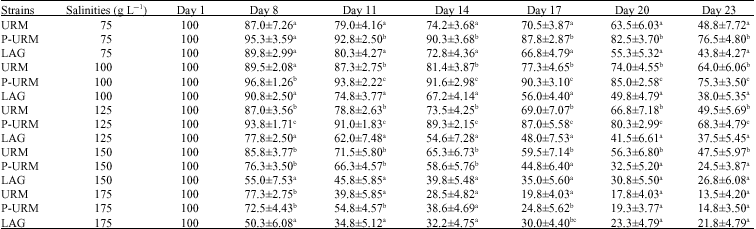 | |
Populations sharing similar letter(s) in each column are not significantly different at respective salinities (p<0.05) | |
| Table 3: | Mean values±SD of total lengths (in mm) of three Artemia populations studied |
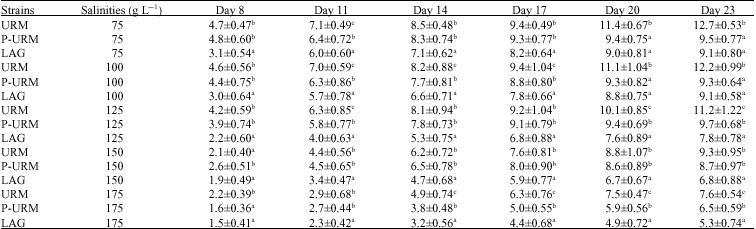 | |
| Populations sharing similar letter(s) (a, b, c) in each column are not significantly different at respective salinities (p<0.05) | |
| Table 4: | Average reproductive and life span characteristics of 3 Artemia populations from Urmia Lake region under the function of salinity |
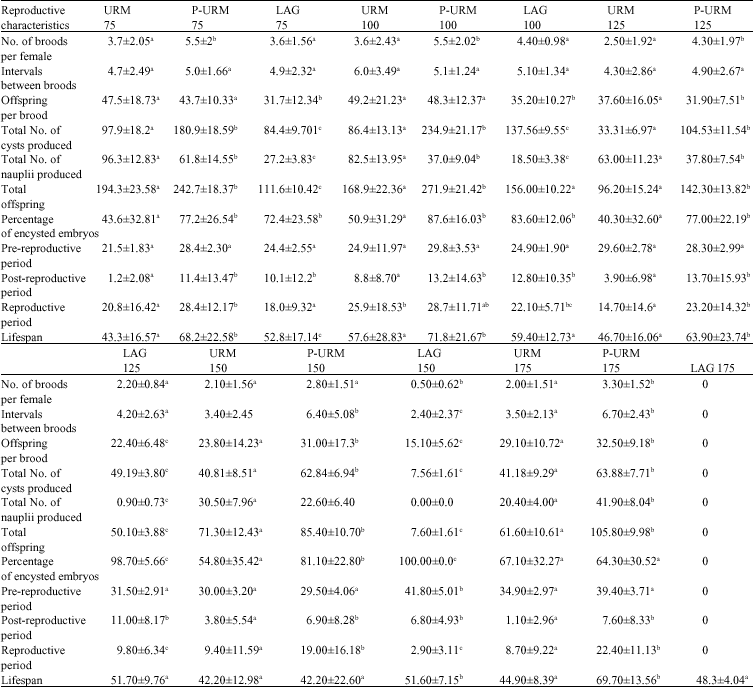 | |
| Populations sharing similar letter(s) a, b, c in each row are not significantly different at respective salinities (p<0.05) | |
in the percentage of encysted embryos and total life span compared to the asexual populations, while P-URM demonstrated significantly higher values in five characters (i.e., number of broods per female, total number of cysts produced, total offspring, reproductive period and total life span compared to URM and LAG (ANOVA, p<0.05, Table 4). At 100 g L-1, URM strain was statistically different from the parthenogenetic strains in four out of eleven reproductive and life span characteristics (i.e., total number of cysts, total number of nauplii, percentage of encysted embryos and post-reproductive period), being significantly higher only in total number of nauplii produced, while P-URM had significantly higher values in number of broods per female, total number of cysts produced, total offspring and total life span compared to URM and LAG strains (ANOVA, p<0.05, Table 4).
At 125 g L-1 URM had significantly higher values only in offspring per brood and total number of nauplii, while P-URM showed significantly higher values in the same 5 characters as in 75 g L-1 (ANOVA, p<0.05, Table 4). At 150 and 125 g L-1, the reproductive and life span pattern were similar. But at 175 g L-1 the asexual P-URM had higher values in almost all studied characters compared to sexual URM, while LAG strain did not reproduce any offspring in this salinity. Comparisons of reproductive and life span variables in the five salinities revealed that most of the characters studied, were statistically different at these salinities. Two variables, percentage of encysted embryos and pre-reproductive period, were significantly affected by salinity, exhibiting a parallel increase with salinity increase up to 150 g L-1 in most cases. Intervals between broods and total life span are hardly affected by salinity but there was a decrease in the rest of the characters as salinity became higher. Highest values for total offspring and offspring per brood in all populations was observed at 75 and 100 g L-1 (Table 4) indicating that the best reproductive performance of these populations was observed at 75 and 100 salinity. Longest life span was observed at 100 g L-1 for all three strains. P-URM lived significantly longer than the other two populations in all salinities.
DISCUSSION
Survival: Although thriving Artemia populations are found in salinities as high as 340 g L-1 in nature (Post and Youssef, 1977), its culture and maintenance in laboratory at salinities higher than 200 g L-1 has always been difficult (Wear and Haslett, 1986; Wear et al., 1986). Browne and Hoopes (1990) reported only 9% survival at 190 g L-1 and no survival at all at 230 g L-1 in a parthenogenetic Artemia from Salin de Giraud (France). Dana and Lenz (1986) studying the bisexual Artemia from Mono Lake, California, USA, found low survival in 159 and 179 g L-1 under laboratory conditions. Triantaphyllidis et al. (1995) reported over 80% mortality of both parthenogenetic Artemia from Tanggu area (China) and A. franciscana at 180 g L-1 at 25°C over a 23 days culture period. On the contrary, they reported above 75% survival for A. franciscana and higher than 50% survival for parthenogenetic Artemia at salinities lower than 100 g L-1.
In the experiments performed by El-Bermawi et al. (2004) on Artemia populations from Egypt, 100% mortality was observed in bisexual A. salina from Wadi El-Natrun in 150 and 200 g L-1 within 17 days, but the high salinity had little effect on the ability of parthenogenetic populations to survive. Total mortality did not occur in our experiments with sexual and asexual strains Artemia from the Urmia Lake and the neighbouring lagoons within the range of 75-175 g L-1 salinities. Browne and Wanigasekera (2000) observed an increase in survival of parthenogenetic Artemia from Margherita di Savoia (Italy) and A. salina when salinity of the culture medium was increased from 60 to 120 g L-1 at 15°C, but this percentage sharply decreased in three other bisexual species (A. sinica, A. franciscana and A. persimilis). Inversely, at 24°C they got completely different results, obtaining significantly higher survival at higher salinity. El-Bermawi et al. (2004) observed similar results with parthenogenetic Artemia populations from Egypt. Contrary to these two findings, Triantaphyllidis et al. (1995) found a steady decrease in survival in both parthenogenetic from Tanggu (China) and bisexual A. franciscana cultured in the range of 60 to 180 g L-1. Triantaphyllidis et al. (1995) reported 70-80% survival for A. franciscana at 60 g L-1, but Browne and Wanigasekera (2000) observed only 16% survival for this species at the same salinity. Vanhaecke et al. (1984) reported high survivorship for A. sinica and A. salina at 60 g L-1, whereas survival was zero for these two species at the same salinity in the experiments performed by Browne and Wanigasekera (2000).
Browne and Wanigasekera (2000) claimed that differences in the culture conditions and intra-species and population-dependent characteristics could be among the reasons for the different results obtained by different researchers. Present experiments are in agreement with the findings of Triantaphyllidis et al. (1995), showing a constant decline in survival when salinity increases from 100 to 170 g L-1. Different findings with different Artemia strains could be an indication for strain-specific adaptation patterns of various Artemia populations to diverse physical, chemical and biotic characteristics of their own biotopes.
Abatzopoulos et al. (2006b) reported very low survival for A. urmiana in salinities of 35 and 50 g L-1. But they found high survival at 100, 140 and 180 g L-1. However, they did not find significant differences in the survival of bisexual A. urmiana in the later elevated salinities.
Present findings are not in agreement with findings of Abatzopoulos et al. (2006b). In present study, we found survival percentages of both sexual and asexual populations that decrease with increasing salinity. According to the results obtained in present study, sexual URM can tolerate higher salinities compared to the two asexual strains. However, high mortality was observed in URM at 175 g L-1, whereas parthenogenetic populations begin experiencing sharp mortality at 150 g L-1. Unlike the Egyptian parthenogenetic populations as reported by El-Bermawi et al. (2004), the Iranian asexual strains had very low survival at high salinities.
Growth: According to Gilchrist (1960), Triantaphyllidis et al. (1995) and El-Bermawi et al. (2004) growth is inversely proportional to salinity. Triantaphyllidis et al. (1995) reported significant differences in the growth of Artemia especially in the parthenogenetic population from Tanggu (China) cultured at different salinities. According to their experiments maximum growth in A. franciscana was observed at 35 g L-1, whereas growth in parthenogenetic Artemia showed no differences in 35, 60 and 100 g L-1. But parthenogenetic Artemia at 180 g L-1 attained only 50% of the length of those at 35, 60 and 100 g L-1. A. franciscana at 180 g L-1 achieved 60% of the length in comparison to animals grown at 35 g L-1. El-Bermawi et al. (2004) did not observe big differences in growth of parthenogenetic and bisexual populations of Artemia from Egypt grown in the laboratory at salinities ranging from 35 to 200 g L-1. Abatzopoulos et al. (2006) found that growth rate of A. urmiana was not affected by the increase of salinity.
The present study confirms that growth rate in Artemia populations from Urmia Lake region (Iran) is inversely proportional to salinity, supporting the findings of Gilchrist (1960), Triantaphyllidis et al. (1995) and El-Bermawi et al. (2004). In our experiment, URM and LAG at 175 g L-1 could attain almost 60% of the total length of those grown at 75 and 100 g L-1, while P-URM at 175 g L-1 achieved almost 75% of the length of 75 g L-1 grown animals. Present results are not in agreement with the findings of Abatzopoulos et al. (2006) and prove that A. urmiana grows best at 75-100 g L-1 and that its growth rate is significantly affected by elevated salinity. It seems that performance of A. urmiana and probably the asexual strains are affected by different sources of brine water used for culture experiments. In present study, we used diluted Lake Urmia water adjusted to 75, 100, 125, 150 and 175 g L-1 throughout the experiment, whereas Abatzopoulos et al. (2006b) used artificially prepared D and K medium of 35, 50, 100, 140 and 180 g L-1 salinity in their experiments. Present results in combination with literature data suggest that adaptation to different salinities and growth rate are species-specific and in addition dependent on the culture conditions.
Reproductive and life span characteristics: A number of investigations have reported on effects of salinity on the reproductive and life span characteristics of Artemia. Gilchrist (1960), Dana and Lenz (1986) and Triantaphyllidis et al. (1995) who worked on A. salina,A. franciscana from Mono lake and Tanggu parthenogenetic Artemia and A. franciscana from San Francisco Bay, respectively, reported that maturation is achieved fastest at salinities lower than 100 g L-1 and much slower above 140 g L-1. Abatzopoulos et al. (2003) reported faster maturity at 50 and 80 g L-1 in comparison to 120 g L-1 for a parthenogenetic Artemia from Megalon Embolon (Greece). Similarly Baxevanis et al. (2004) reported early maturation at 35 g L-1 in three parthenogenetic populations and at 80 g L-1 in the bisexual A. salina from Lake of Wadi El-Natrun, all from Egypt. It was found that this bisexual Artemia died before attaining maturity at 150 and 200 g L-1. But Browne and Wanigasekera (2000) who performed the experiments at various combinations of temperature and salinity with five Artemia populations (one parthenogenetic and four bisexual) reported parthenogenetic Artemia from Margherita di Savoia (Italy) as a niche specialist attaining maturity and reproducing only at salinities higher than 120 g L-1 at 24°C. This Artemia was not able to reproduce at 60 g L-1 at 15°C or 30°C. According to their findings maturation time in all four bisexual populations was more temperature-dependent than salinity-dependent. Within different temperature treatments at lower salinity, maturity was achieved earlier than at higher salinities, except in A. persimilis which had the shortest maturation time (9.7 days) at 30°C at 180 g L-1 (Browne and Wanigasekera, 2000) Triantaphyllidis et al. (1995) did not find significant differences in the reproductive characteristics (offspring per brood, broods per female, offspring per female per day, days between broods, total offspring per female and percentage of encystment) in Tanggu parthenogenetic Artemia and A. franciscana from San Francisco Bay at interpopulation level, but there were significant differences at intrapopulation level at salinities below 100 g L-1. Abatzopoulos et al. (2003) studying an Artemia clone from Megalon Embolon observed significant differences in all reproductive characteristics except for number of broods in all three salinities (50, 80, 120 g L-1). Baxevanis et al. (2004) reported significant differences in most of the reproductive parameters between the bisexual and parthenogenetic populations he studied at 35, 80 and 120 g L-1. Moreover they observed statistically significant differences between the inland and coastal parthenogenetic populations of Artemia from Egypt, but no differences were evident between the two coastal strains. Browne and Wanigasekera (2000) reported that all five populations in their study had highest reproduction period and peak production at 24°C at either 120 or 180 g L-1. According to their findings Old World species (A. sinica and A. salina) and the parthenogenetic population are more limited by temperature and salinity for reproduction, whereas New World species (A. franciscana and A. persimilis) are euryhaline and eurythermal being able to reproduce more successfully at more diverse salinity-temperature combinations. But present results indicate that A. urmiana and the two asexual populations from Iran are able to reproduce very well pat all salinities below 150 g L-1 and peak production was observed at 75 and 100 g L-1.
In this study, which was carried out at five different salinities ranging from 75 to 175 g L-1 at 27±1°C, all three populations did produce offspring in all salinities, except for the parthenogenetic Artemia from lagoons, which was not reproductive at 175 g L-1. Present results are in agreement with findings of Gilchrist (1960), Dana and Lenz (1986), Triantaphyllidis et al. (1995), Abatzopoulos et al. (2003) and Baxevanis et al. (2004) conforming that reproductive and life span characteristics are inversely affected by salinity. Present results also proved that early maturation is achieved by sexual URM and the two asexual populations from Lake Urmia and neighboring lagoons in low salinities. Unlike findings of Triantaphyllidis et al. (1995) with Tanggu (China) parthenogenetic Artemia and A. franciscana from San Francisco Bay, we found significant differences in most of the reproductive and life span characteristics of URM and two asexual populations from Iran both at inter and intrapopulation levels in all salinities. In accordance with findings of Baxevanis et al. (2004) on Egyptian sexual and asexual populations, we also found significant differences in reproductive and life span characteristics of Iranian sexual and parthenogenetic strains under function of different salinities.
Abatzopoulos et al. (2006b) did not find significant differences in five out of six reproductive traits and two out of four life span characteristics of A. urmiana in different salinities. According to them A. urmiana cultured at 140 and 180 g L-1 showed a significantly longer reproductive period and total life span than those at 100 g L-1. Same study also revealed that A. urmiana has higher tendency for encystment with salinity increase. Baxevanis et al. (2004) observed that the salinity increase was positively correlated with the number of produced encysted embryos in Egyptian sexual and asexual Artemia populations. Unlike findings of Abatzopoulos et al. (2006b), we observed significant differences in 3 major reproductive characteristics i.e., total offspring, number of cysts and number of nauplii produced in A. urmiana cultured at different salinities, higher offspring production was observed at lower salinities (75 and 100 g L-1). But our results support findings of Abatzopoulos et al. (2006b) and Baxevanis et al. (2004) on percentage of encystment by Artemia populations at elevated salinities, although in this study total cyst production decreased significantly at 150 and 180 g L-1 compared to 75 and 100 g L-1.
The present study supports findings by many studies on the negative impact of salinity values above 120-140 g L-1 on reproductive and life span characteristics in many other Artemia species or strains (Vanhaecke et al., 1984; Wear and Haslett, 1986; Triantaphyllidis et al., 1995; Browne and Wanigasekera, 2000; Baxevanis and Abatzopoulos, 2004; Baxevanis et al., 2004). In accordance with previous laboratory investigations on several Artemia species (Browne et al., 1991; Triantaphyllidis et al., 1995; Baxevanis et al., 2004), it was found that the optimal range for growth, survival and reproduction of Iranian sexual and asexual strains of Artemia from Lake Urmia region lies between 75 and 125 g L-1.
Although salinity was usually very high, ranging from 250-300 g L-1 during the last ten years in most parts of the Lake Urmia (Agh, 2006; Agh et al., 2007), considerably high Artemia biomass concentrations could be seen at limited areas in the lake during spring and summer. It should be remembered that numerous quantities of Artemia cysts hatch in Lake Urmia during spring and summer in areas with low salinities. Therefore it could be assumed that the newly hatched nauplii go through an adaptation period before migrating to deeper parts of the lake where salinity never dropped below 250 g L-1 since 1998. However there are no accurate data on mortality rates of Artemia in Lake Urmia. Apparently the total number of adults in the lake is much less than the total number of nauplii hatching from cysts in spring and summer (Agh, unpublished data). This would mean that even in a natural biotope only limited number of animals that can withstand high salinity levels can survive and the rest die off. This could be a logic explanation for thriving Artemia populations at very high salinities in nature. This phenomenon could also help us to understand why culture and maintenance of Artemia in laboratory at salinities higher than 200 g L-1 has always been difficult and why very high mortality occurs under function of high salinities in laboratory culture of Artemia. Thus it could be concluded that, despite extraordinary tolerance levels of Artemia to high salinities, it seems that salinity is the crucial limiting factor in controlling the population density of Artemia both in nature and in the laboratory.
ACKNOWLEDGMENTS
This study was carried out with financial support of the Ministry of Science, Research and Technology, Islamic Republic of Iran. We also acknowledge the EU INCO-DEV Concerted Action project Artemia Biodiversity (ICA4-CT-2001-10020), that partially supported this research. Technical assistance by the staff of the Artemia and Aquatic Animals Research Center, Urmia University and Artemia Reference Center, Ghent University are also greatly appreciated.
REFERENCES
- Abatzopoulos, T.J., N. El-Bermawi, C. Vasdekis, A.D. Baxevanis and P. Sorgeloos, 2003. Effects of salinity and temperature on reproductive and life span characteristics of clonal Artemia.(International Study on Artemia. LXVI). Hydrobiologia, 492: 191-199.
CrossRefDirect Link - Abatzopoulos, T.J., N. Agh, G. Van-Stappen, S.M. Razavi-Rouhani and P. Sorgeloos, 2006. Artemia sites in Iran. J. Mar. Biol. Assoc. UK., 86: 299-307.
CrossRefDirect Link - Abatzopoulos, T.J., A.D. Baxevanis, G.V. Triantaphyllidis, G. Criel, E.L. Pador, G. Van Stappen and P. Sorgeloos, 2006. Quality evaluation of Artemia urmiana Gunther (Urmia Lake, Iran) with special emphasis on its particular cyst characteristics (International Study on Artemia LXIX). Aquaculture, 254: 442-454.
CrossRefDirect Link - Agh, N., T.J. Abatzopoulos, I. Kappas, G. Van Stappen, S.M.R. Rouhani and P. Sorgeloos, 2007. Coexistence of sexual and parthenogenetic Artemia populations in lake Urmia and neighbouring Lagoons. Int. Rev. Hydrobiol., 1: 48-60.
CrossRefDirect Link - Baxevanis, A.D. and T.J. Abatzopoulos, 2004. The phenotypic response of ME2 (M. Embolon, Greece) Artemia clone to salinity and temperature. J. Biol. Res., 1: 107-114.
Direct Link - Baxevanis, A.D., N. El-Bermawi, T.J. Abatzopoulos and P. Sorgeloos, 2004. Salinity effects on maturation, reproductive and life span characteristics of tour Egyptian Artemia populations (International Study on Artemia, LXVIII). Hydrobiologia, 513: 87-100.
Direct Link - Browne, R.A., L.E. Davis and S.E. Sallee, 1988. Effects of temperature and relative fitness of sexual and asexual brine shrimp Artemia. J. Exp. Mar. Biol. Ecol., 124: 1-20.
CrossRefDirect Link - Browne, R.A. and C.W. Hoopes, 1990. Genotype diversity and selection in asexual brine shrimp (Artemia). Evolution, 44: 1035-1051.
CrossRefDirect Link - Browne, R.A. and G. Wanigasekera, 2000. Combined effects of salinity and temperature on survival and reproduction of five species of Artemia. J. Exp. Mar. Biol. Ecol., 244: 29-44.
Direct Link - Browne, R.A., V. Moller, V.E. Forbes and M.H. Depledge, 2002. Estimating genetic and environmental components of variance using sexual and clonal Artemia. J. Exp. Mar. Biol. Ecol., 267: 107-119.
Direct Link - Clark, L.S. and S.T. Bowen, 1976. The genetics of Artemia salina. VII. Reproductive isolation. J. Heredity, 67: 385-388.
PubMedDirect Link - Cole, G.A. and R.J. Brown, 1967. The chemistry of Artemia habitats. Ecology, 48: 858-861.
CrossRefDirect Link - Dana, G.L. and P.H. Lenz, 1986. Effects of increasing salinity on an Artemia population from Mono lake, California. Oecologia, 68: 428-436.
CrossRefDirect Link - El-Bermawi, N., A.D. Baxevanis, T.J. Abatzopoulos, G. Van Stappen and P. Sorgeloos, 2004. Salinity effects on survival, growth and morphometry of four Egyptian Artemia populations (International Study on Artemia. LXVII). Hydrobiologia, 523: 175-188.
Direct Link - Gilchrist, B.M., 1960. Growth and form of the brine shrimp Artemia salina (L.). Proc. Zool. Soc. Lond., 134: 221-235.
Direct Link - Kappas, I., T.J. Abatzopoulos, N.V. Hoa, P. Sorgeloos and J.A. Beardmore, 2004. Genetic and reproductive differentiation of Artemia franciscana in a new environment. Mar. Biol., 146: 103-117.
Direct Link - Post, F.J. and N.N. Youssef, 1977. A procaryotic intracellular symbiont of the great salt lake brine shrimp Artemia salina (L.). Can. J. Microbiol., 23: 1232-1236.
CrossRefDirect Link - Sorgeloos, P., 1997. Lake Urmia cooperation project-contract item a report on the determination and identification of biological characteristics of Artemia urmiana for application in aquaculture. Faculty of Agriculture and Applied Biological Science, Laboratory of Aquaculture and Artemia Reference Center, Ghent University, Belgium, pp: 6-16.
- Triantaphyllidis, G.V., K. Poulopoulou, T.J. Abatzopoulos, C.A.P. Perez and P. Sorgeloos, 1995. International study on Artemia XLIX. Salinity effects on survival, maturity, growth, biometrics, reproductive and lifespan characteristics of a bisexual and a parthenogenetic population of Artemia. Hydrobiologia, 302: 215-227.
CrossRefDirect Link - Triantaphyllidis, G.V., G.R.J. Criel, T.J. Abatzopoulos, K. Thomas, J. Peleman, J.A. Beardmore and P. Sorgeloos, 1997. International study on Artemia. LVII. Morphological and molecular characters suggest conspecificity of all bisexual European and North African Artemia populations. Mar. Biol., 129: 477-487.
CrossRefDirect Link - Triantaphyllidis, G.V., G.R. Criel, T.J. Abatzopoulos and P. Sorgeloos, 1997. International study on Artemia. LIII. Morphological study of Artemia with emphasis to Old World strains. I. Bisexual populations. Hydrobiologia, 357: 139-153.
CrossRefDirect Link - Vanhaecke, P., S.E. Siddall and P. Sorgeloos, 1984. International study on Artemia. XXXII. Combined effects of temperature and salinity on the survival of Artemia of various geographical origin. J. Exp. Mar. Biol. Ecol., 80: 259-275.
CrossRefDirect Link - Vanhaecke, P., W. Tackaret and P. Sorgeloos, 1987. The biogeography of Artemia: An updated review. Artemia Res. Applic., 1: 129-155.
Direct Link - Van Stappen, G., G. Fayazi and P. Sorgeloos, 2001. International study on Artemia LXIII. Field study of the Artemia urmiana (Gunther, 1890) population in Lake Urmiah, Iran. Hydrobiologia, 466: 133-143.
CrossRefDirect Link - Browne, R.A., S.E. Salee, D.S. Grosch, W.O. Segreti and S.M. Purser, 1984. Partitioning genetics and environmental components of reproduction and life span in Artemia. Ecology, 65: 949-969.
CrossRefDirect Link