
Fatma A. Elhumeed
Department of Zoology, Faculty of Education of Girls, Saudi Arabia
Wafaa B. Yousif
Department of Zoology, Faculty of Science, Alexandria, Egypt
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 2 | Page No.: 153-163







ABSTRACT
The present study was an investigation of the effect of the contraceptive drug, Nordette, on the stomach of the mouse when administered daily at a recommended therapeutic dose rate of 0.0026 mg kg-1 for 30 days. Extensive light and electron microscopic changes were noticed. The drug caused enlargement in the all types of cells. The oxyntic cells appeared hypertrophied with irregular cell boundaries, enlarged nuclei and faintly stained cytoplasm. Their cytoplasm contained irregularly distributed mitochondria with dense matrix, decreased rER, obviously increased sER, disorganized intracellular canaliculi and some lysosomes. The peptic cells appeared enlarged and contained hypertrophied rough endoplasmic reticulum and an increase amount of ribosomes and secretory granules. There was an increase in the amount of the secretory granules in the lumen of the gastric gland. The mucus cells at the upper region of the gastric gland were greatly decreased. Smooth muscle fibers showed enlargement and degeneration. The submucosa and lamina propria showed vacuolation. The most pathological effects were restricted to the obvious decrease of the lymphoid cells in the submucosa and lamina propria. Dilatation and congestion of the blood vessels and blood capillaries were noticed. Blood capillaries lined by enlarged endothelial cells containing enlarged heterochromatic nuclei.
PDF Abstract XML References Citation
How to cite this article
Fatma A. Elhumeed and Wafaa B. Yousif, 2008. Light and Electron Microscopic Studies on the Effect of a Contraceptive Drug on the Stomach of Mouse. Pakistan Journal of Biological Sciences, 11: 153-163.
DOI: 10.3923/pjbs.2008.153.163
URL: https://scialert.net/abstract/?doi=pjbs.2008.153.163
DOI: 10.3923/pjbs.2008.153.163
URL: https://scialert.net/abstract/?doi=pjbs.2008.153.163
INTRODUCTION
Estrogen has various physiological functions such as growth and differentiation, via not only endocrine but autocrine or paracrine fashion (Simpson et al., 1999). Though the ovary is a major source of systemic estrogen, estrogen is produced also in extraovarian tissues including, adipose tissue (Ackerman et al., 1981), skin (Leshin et al., 1981), osteoblasts (Purohit et al., 1992), testis (Brodie and Inkster, 1993), brain (Lephart, 1996), vascular smooth muscle cells (Harada et al., 1999), T cells in the spleen (Samy et al., 2001). These extraovarian tissues, however probably do not produce a sufficient quantity of estrogen to affect the circulatory level. Recently, Ueyama et al. (2006) stated that under normal physiological condition a large quantity of estrogen equivalent to ovary is secreted from the unidentified region of the gastrointestinal organs into the portal vein and most of it is then trapped in the liver, but in the case of increased portal systemic shunting it overflows into the systemic circulation resulting in the elevation of systemic estrogen concentration to cause estrogen-excess signs associated with liver cirrhosis. Patients with liver cirrhosis often show an increase of serum estradiol (E2)/testosterone ratio, which is supposedly caused by an abnormal estrogen metabolism in the liver. Gene expression of aromatase, estrogen synthetase, was demonstrated in the gastric mucosa of male and female adult rats, Parietal cells exhibited strong signals for aromatase mRNA and immunoreactive protein by in situ hybridization histochemistry and immunohistochemistry. These findings indicate that gastric parietal cells play a potent endocrine role in secreting estrogen that may function as a regulator of the gastro-hepatic axis. These data showed that the gastric mucosa can produce a large quantity of estrogen equivalent to that in the ovary (Ueyama et al., 2006). Previous studies showed that experimental portal-systemic shunting in adult male rats resulted in a marked increase of systemic estrogen concentration (Farrell et al., 1988). Matsuyama et al. (2002) studied expression Estrogen Receptor (ER) beta in human 29 stomach adenocarcinoma and stated that the effects of estrogen in stomach cancer, as well as those in normal stomach may be mediated by ER beta.
Jiang et al. (2006) stated that exposure to estrogens is associated with an increase risk of developing breast, cervical and liver cancer. Estrogens strongly induce the human granzyme B inhibitor, proteinase inhibitor 9 (PI9). Cytotoxic T lymphocytes (CTLs) and Natural Killer (NK) cells use the granzyme pathway to induce apoptosis of target cells. Yousif (1997) and Yousefi et al. (1988) reported that sex hormones including estrogen, progesterone and testosterone are known to have adverse effect on immune system and particularly on the proliferative response.
Humans are routinely exposed to bisphenol-A (BPA), an estrogenic compound that leaches from dental material, food and beverage container and other plastic consumer products. Vandenberg et al. (2007) showed that BPA induced adverse effect in mammary glands development in the embryo. Correspondence to Stallard et al. (2000), hormone replacement therapy is being used increasingly. Although it is known that the risk of developing breast cancer is slightly increased with long term use (Gambrell, 1992).
Li et al. (2006) studied how the estrogen modulates the rigidity of the cytoskeleton in the epithelial cells; estrogen depletion decreased and treatment with 17β-estradiol increased deformability of cervical-vaginal epithelial cells. Estrogen also induced redistribution of nonmuscle myosin II-B(NMM-II-B); lesser interaction of NMM-II-B with actin; increased phosphorylation of NMM-II-B heavy chains and decreased filamentation of NMM-II-B in vitro. Similarly, Gorodeski (2007) tested the hypothesis that estrogen control epithelial paracellular resistance through modulation of myosin and stated that estrogen modulates nonmuscular myosin (NMM)-II-B, the main component of cortical actomyosin in human epithelial cervical cells.
This study aims to examine the histopathological and the ultrastructural changes in the stomach of mice treated with oral therapeutic doses of a contraceptive drug.
MATERIALS AND METHODS
The experiment was conducted at the Faculty of Science laboratory, Alexandria University, Egypt. Female Swiss mice (Mus musculus) (from the animal house of the High Health Institute) of 8 weeks old and 20-22 g in weight were used in this study. Food and water were provided ad libitum. Nordette tablets (Levonorgestrel 0.15 mg and Ethyl estradiol 0.03 mg, manufactured by The Nile Company for Pharmaceutical, Cairo) were uniformly suspended in distilled water. The animals were treated daily intragastrically via a bent stainless steel feeding tube with a therapeutic dose of 0.0026 mg kg-1 of Nordette for 30 days. The dose used in the present study was the recommended therapeutic dose. A group of female mice of similar age and weight was as control and received no treatment. Small slices of stomach were immediately fixed in 2.5% glutaraldehyde buffered with phosphate buffer (pH 7.6) for 1 h and post fixed in 1% osmium tetraoxide (in the same buffer) for 1-2 h at 4°C. Specimens were dehydrated through graded series of ethyl alcohol and embedded in Araldite-Epon mixture. Semithin (1 μm) and ultrathin (50 nm) sections from selected areas were cut with a glass knife on LKB ultramicrotome. Semithin section were stained with Toluidine Blue (TB) and examined with light microscopy, while ultrathin section were double stained with uranyl acetate and lead citrate and examined by Jeol 100 CX Electron.
RESULTS
Light microscopy of control stomach: The gastric mucosa consists of a surface epithelium that invaginated to varying extend into the lamina propria, forming pits. Emptying into the gastric pits are long straight, branched tubular glands. The lamina propria of the stomach, which extend between the glands is composed of loose connective tissue interspersed with blood and lymph vessels, smooth muscle fibers and lymphoid cells (Fig. 1- 4). The lamina propria supported the gastric glands (Fig. 1- 4). The submucosa is relatively loose and distensible and the larger blood vessels and a lymphoid aggregate lie in it (Fig. 1, 2). Separating the mucosa from the underlying submucosa is a continuous layer of smooth muscle, muscularis mucosa (Fig. 2). The stomach has a well developed muscularis mucosa. Muscularis mucosa lying immediately beneath the base of gastric glands clearly demarcating them from the underlying submucosa (Fig. 2). Stips of muscularis mucosa may closely associated with gastric glands and the contraction
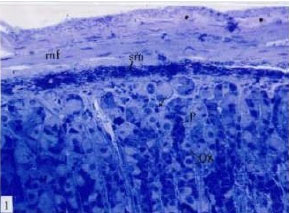 | |
Fig. 1: | Semithin section of stomach of control mouse, showing oxyntic cells (Ox) peripherally in the gland, small peptic cells (P), smooth muscle fibers (mf), lymphoid cells in submucosa (sm) and lamina propria (arrow), nerve plexuses (*) toluidine blue (TB) (x400) |
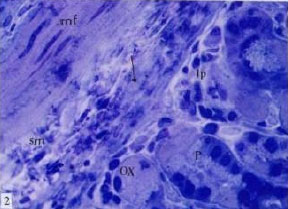 | |
Fig. 2: | Semithin section of stomach of control mouse, showing peptic cells (P) with basal nucleus and apical refractile secretory granules, oxyntic cells (Ox), lymphoid cells in submucosa (sm) and lamina propria (lp), muscularis mucosa (arrow) TB (x1000) |
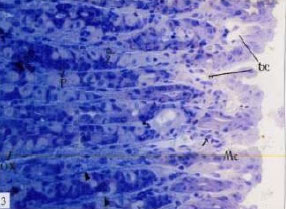 | |
Fig. 3: | Semithin section of stomach of control mouse, showing blood capillaries (bc) under thick, well developed mucus cell (Mc) layer, oxyntic cells (Ox), peptic cells (P), lymphoid cells (arrow), smooth muscle fibers (head arrow) TB (x400) |
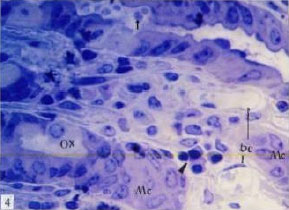 | |
Fig. 4: | Semithin section of stomach of control mouse, showing many large blood capillaries (bc) under surface mucus cell (Mc), oxyntic cells (Ox), few secretory granules in gastric gland (arrow), lymphoid cells (head arrow) TB (x1000) |
of these smooth muscle cells may help gastric glands express their secretion (Fig. 3). The muscularis consists of an outer thin longitudinal and inner thick circular smooth muscle layers, it is composed of bundles of smooth muscle fibers held together by loose connective tissue. Nerve plexuses are seen between the outer and inner muscle layers (Fig. 1).
The gastric mucosal coat appear thick due to the presence of gastric gland. The gastric glands contain a mixed population of endocrine cells and different type of exocrine cells, mucus cells, oxyntic cells and peptic cells.
Mucous-secreting cells:The epithelium covering the surface and lining the pit is a simple columnar mucus cells, all the cells secrete mucin (Fig. 3, 4). The Mucous surface cells are the most superficial cells in the gland. They extend a short distance into the gastric gland. Mucus-secreting cells of the stomach are not of the goblet form found in other parts of the body. Other mucus secreting cells in the necks and ischmus portions of the gastric glands, called neck mucous cells they secrete a less viscous mucus which may protect the gland duct from autodigestion. Mucus neck cells are slightly different cells, their mucus has different chemical properties from the mucus secreted by mucous surface cells. Neck mucous cells are smaller and less regular in shape than surface mucous cells, mainly because they are compressed and distorted by adjacent cells. They have basal nucleus and finely granular cytoplasm due to the presence of small mucin vacuoles (Fig. 3, 4).
Oxyntic (parietal) cells: Oxyntic cells are distributed along the length of the glands but tend to be most heavily concentrated in the upper and middle portions of gastric glands, where they are mixed with peptic cells and stem cells. They are larger than the other cells of the gastric glands. They can be recognized by their characteristic fried egg appearance. They are located peripherally in the gland so their broad bases appear to bulge into the underlying lamina propria. Oxyntic cells vary slightly in their shape and size (Fig. 1- 4). The much larger oxyntic cells are pyramidal, oval, or rounded cells, have centrally located large spherical nucleus of low density and pale eosinophilic cytoplasm which often appears vacuolated, particularly around the nucleus. Their attachment to the basal membrane is broad, but their luminal aspect is narrow being compressed between adjacent cells (Fig. 1- 4). The eosinophilic cytoplasm is due to the numerous mitochondria (seen by electron microscope) which are a feature of highly metabolically-active cells.
Peptic (chief) cells: Peptic cells are the principle cell types in the basal third of the gastric glands. These cells appear small, irregular pyramidal, or cuboidal in shape. Peptic cells are recognized by their condensed basally located large nuclei and strongly basophilic basal cytoplasm. They contain apical refractile large cytoplasmic granules (Fig. 1-3).
Stem cells: stem cells are the precursor cells of all epithelial cells of the gastric mucosa. They are small cells with oval basal large nuclei and small amount of cytoplasm and show no cytoplasmic specialization when completely undifferentiated. Occasionally, the enteroendocrine cells can be identify in the section, characterized by their triangular shape.
Because of increase of the array of the subsurface capillaries, it seems certain that virtually all surface mucus cells will be in close proximity to this capillary net work (Fig. 3, 4). Progressing towards the gastric lumen the amount of the blood capillaries increased and become obviously enlarged, there was a notable decrease in the amount of connective tissue between the basal lamina of the endothelium and that of the adjacent gastric gland mucus cells (Fig. 4).
Electron microscopy of control stomach:
Oxyntic cells: Oxyntic cell nuclei contain relatively few moderately electron-dense hetrochromatin, many nuclear pores and obvious nucleoli (Fig. 5). They have an elaborate system of apical surface invaginations called intracellular canaliculi, consist of a branching system of passages within the cell, opening at the free surface of the cell to communicate either directly or via an intercellular canaliculus with the lumen of the gland i.e., the luminal plasma membrane forms deep, branching canaliculi which extend throughout the cytoplasm and between adjacent cells, numerous short microvilli project into the lumina of intracellular canaliculi greatly increasing the surface area. The canaliculi are closely associated with mitochondria and an abundance of smooth endoplasmic reticulum. These features facilitate the active transport of hydrogen ions across a large concentration gradient in the resting cell. In survey micrographs the ovoid, large mitochondria are the dominant feature of the cytoplasm The cytoplasm possess a great number of mitochondria with closely packed cristae. The mitochondria appear to be separated into peripheral group and a central perinuclear group by a ring-like lighter zone of the cytoplasm containing the intracellular canaliculi. The cytoplasm of the oxyntic cells have a discrete, small Golgi apparatus near the cell base and some rough endoplasmic reticulum (Fig. 5).
Peptic cells: Peptic cells predominate in the lower region of the gastric glands. Their basophilia is due to the abundant rough endoplasmic reticulum. Peptic cells have
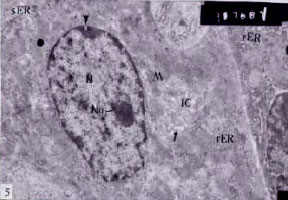 | |
Fig. 5: | Ultrathin section of stomach of control mouse, showing oxyntic cell contain nucleus (N) with moderately dense heterochromatin, nucleolus (Nu), nuclear pore (head arrow), intracellular canaliculi (IC), microvilli (arrow), mitochondria (M), rER, few sER, part of peptic cell with moderate amount of rER (x7.500) |
 | |
Fig. 6: | Ultrathin section of stomach of control mouse, showing thin smooth muscle fibers contain contractile protein, nucleus (N), mitochondria (M), thin and regular endmycium (arrow) (x5000) |
large basal nucleus containing large nucleolus and moderate amount of heterochromatin. They contain large membrane bounded cytoplasmic secretory granules and a rich rough endoplasmic reticulum, a prominent Golgi apparatus and a usual number of mitochondria (Fig. 5). Smooth muscle fibers were thin, containing elongated nucleus with moderately electron-dense heterochromatin, contractile proteins, some mitochondria and thin endmycium (Fig. 6).
Light and electron microscopy of Nordette-treated stomach: The changes induced by Nordette administration were confined to gastric gland cells, smooth muscle fibers, connective tissue. The connective tissue manifested obvious symptoms of lesions. These
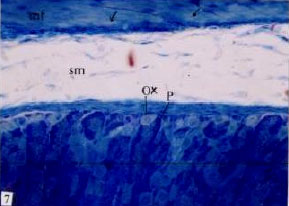 | |
Fig. 7: | Semithin section stomach of Nordette-treated mouse, showing submucosa (sm) devoid of lymphoid cells, oxyntic cells (Ox), peptic cells (P), degeneration (arrow) in smooth muscle fibers (mf). TB (x400) |
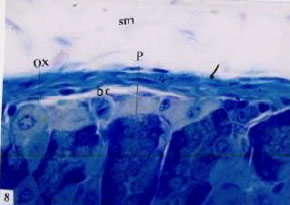 | |
Fig. 8: | Higher magnification of the above micrograph, showing peptic cells (P) which highly basophilic basally, an increase in the apical refractile secretory granules, blood capillaries (bc) muscularis mucosa (arrow). TB (x1000) |
pathological symptoms included marked vacuolation of the submucosa and marked decrease in their cellularity. The most pathological effects were restricted to the absence of the lymphoid cells in the submucosa and lamina propria (Fig. 7- 10). Although, there were many lymphoid cells in the submucosa and lamina propria of the stomach of control animals the submucosa and lamina propria of the treated stomach were nearly devoid of lymphoid cells. Smooth muscle layer of the treated stomach showed an increased in their thickness. The muscle fibers appeared enlarged and contained enlarged nuclei and some of the muscle fibers showed obvious degeneration (Fig. 9, 10). The light microscopic examination revealed an obvious change in the blood vessels and blood capillaries including dilatation and congestion and lined by enlarged endothelial cells
 | |
Fig. 9: | Semithin section of stomach of Nordette-treated mouse, showing degeneration in smooth muscle fibers (arrow), nerve plexuses (*), vacuolated submucosa (sm). TB (x400) |
 | |
Fig. 10: | Semithin section of stomach of Nordette-treated mouse, showing degenerated smooth muscle fibers (arrow), enlarged nucleus (N), submucosa (sm). TB (x1000) |
(Fig. 11). The histopathological changes of the stomach oxyntic cells of treated animals showed some changes. Oxyntic cells appeared hypertrophied with enlarged nuclei and faintly stained cytoplasm (Fig. 11- 13). Some cells possess ill definite cell boundaries. Some oxyntic cells and peptic cells appeared irregular in outline. Peptic cells appeared hypertrophid, with highly basophilic basal cytoplasm and an increase in the amount of the refractile secretory granules in the apical cytoplasm (Fig. 8). The gastric gland lumen contained an increase amount of the secretory granules especially in the upper region of the gland (Fig. 11- 13). The mucus cell layer at the apical region of the gastric gland was greatly decreased (compare Fig. 3 of the control and Fig. 11 of the treated).
Significant ultrastructural changes were revealed by electron microscope. Several unusual structural features were observed in oxyntic cells including some nuclei were
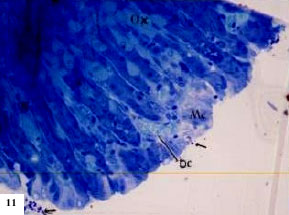 | |
| Fig. 11: | Semithin section of stomach of Nordette-treated mouse, showing oxyntic cells (Ox), obvious decrease in mucus cells (Mc), dilated and congested blood capillaries (bc), secretory granules at the gastric pit (arrow) TB (x400) |
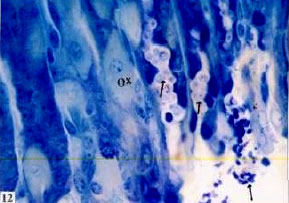 | |
| Fig. 12: | Semithin section of stomach of Nordette-treated mouse, showing oxyntic cells (Ox), increase of secretory granules (arrow) in the gland lumen and discharge of it TB (x1000) |
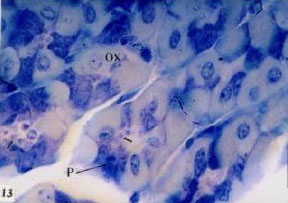 | |
| Fig. 13: | Semithin section of stomach of Nordette-treated mouse, showing transverse section of the gastric gland, oxyntic cells (Ox), peptic cells (P), increase of secretory granules (arrow) in the gland lumen. TB (x1000) |
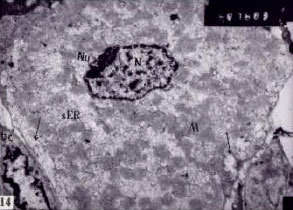 | |
| Fig. 14: | Ultrathin section of stomach of Nordette-treated control mouse, showing oxyntic cell with irregular cell boundaries contain irregular nucleus (N) with highly electron-dense heterochromatin, peripheral dense nucleolus (Nu), hypertrophied sER, increased, mitochondria (M) with dense matrix, peripheral cytoplasmic vacuoles (arrow) underneath a very thin diffusion distances between capillaries (bc) and adjacent oxyntic cells (x5000) |
| |
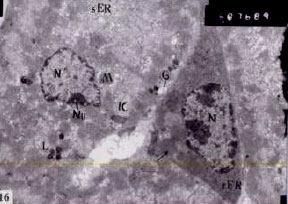
| Fig. 15: | Ultrathin section of stomach of Nordette-treated mouse, showing oxyntic cell, nucleus (N), nucleolus (Nu), mitochondria (M) with dense matrix, lysosome (L), intracellular canaliculi (IC) with irregular microvilli, Note endothelial cells (E) with heterochromatic nucleus, vacuolated basal lamina (arrow), (x7.500) |
irregular, with increase in nuclear pores, contained dense heterochromatin and peripherally located dense nucleoli (Fig. 14). The mitochondria were irregularly distributed in the cytoplasm and not appeared to be separated into peripheral group and a central perinuclear group. The mitochondria increased and in some cells appeared smaller in size and contained denser matrix. The most characteristic features in the oxyntic cells were the pronounced increased and dilatation of smooth
 | |
| Fig. 16: | Ultrathin section of stomach of Nordette-treated mouse, showing oxyntic cell, nucleus (N) with peripheral nucleolus (Nu), mitochondria (M), sER intracellular canaliculi (IC), hypertrophied Golgi apparatus (G), lysosomes (L). peptic cell contain increased rER and large secretory granules (arrow) (x5000) |
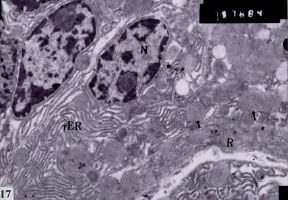 | |
| Fig. 17: | Ultrathin section of stomach of Nordette-treated mouse, showing hypertrophied peptic cells, nucleus (N), increase in rER, ribosomes (R) and secretory granules (arrow) (x7.500) |
endoplasmic reticulum and an obvious decrease of the rough endoplasmic reticulum (Fig. 14). The cytoplasm contained some lysosomes (Fig. 15, 16); lysosomes were not noticed in the oxyntic cells of the control animals. The Golgi apparatus showed hypertrophy and increased activity (Fig. 15, 16). The intracellular canaliculi of some cells were not clearly noticed as in the case of the control and in other cells appeared increased (Fig. 16). The cytoplasm of oxyntic cells beside the blood capillaries contained cytoplasmic vacuoles (Fig. 15). The peptic cells of the treated stomach appeared enlarged and contained hypertrophied rough endoplasmic reticulum and an increase amount of secretory granules in the apical cytoplasm (Fig. 16, 17). Blood capillaries lined by enlarged endothelial cells containing heterochromatic, enlarged nuclei. Endothelial cell cytoplasm contained
 | |
| Fig. 18: | Ultrathin section of stomach of Nordette-treated mouse, showing hypertrophied smooth muscle fibers, hypertrophied heterochromatic nucleus (N) with three electron-dense nucleoli (Nu), many ribosomes in degenerated muscle fibers (arrow) (x5000) |
lysosomes and ribosomes (Fig. 14, 15). The smooth muscle fibers appeared enlarged and contained enlarged heterochromatic nuclei which contained many nucleoli. Some of the muscle fibers showed degeneration in their contractile proteins and contained numerous ribosomes (Fig. 18).
DISCUSSION
In the present study, the administration of Nordette induced histopathological and ultrastructural alterations into stomach of mice. These pathological symptoms included alteration in gastric gland cells, blood vessels, lymphoid cells and smooth muscle fibers.
No work has been reported on possible histopathological effects of the contraceptive drug on the stomach. Many previous reports demonstrated histopathological effects of the contraceptive drug on some other body organs including immune organs (Yousif, 1997), uterus (Hassanein, 2000) and pancreas (El Rawi and Yousif, 2006). Results of the present work revealed that the histopathological changes of the stomach oxyntic cells of treated animals included increase in size, weaker stainability of the cytoplasm. Ultrastructural alterations included an irregular distribution of the mitochondria in the cytoplasm, an obvious decrease of rough endoplasmic reticulum and an increased and dilatation of smooth endoplasmic reticulum, the Golgi apparatus showed hypertrophy and increased activity and the cytoplasm contained some lysosomes, the cytoplasm of oxyntic cells beside the blood capillaries contained cytoplasmic vacuoles. The intracellular canaliculi of some cells were not clearly noticed as in the case of the control. In the present study, peptic cells showed an increase in rER, ribosomes, zymogen granules. Similarly, Hassanein (2000) stated that the administration of oestradiol benzoate stimulated the secretory activity of the uterine luminal epithelial cells as evidenced by the development of rough endoplasmic reticulum, Golgi complexes and the abundance of secretory granules and provokes other alterations including mitochondria were large and swollen, most of their cristae got destructed and many lysosomes appeared in the cytoplasm. Also the administration of Nordette caused increase in rER, ribosomes, zymogen granules in the pancreatic acinar cells (El Rawi and Yousif, 2006). In previous report, Yousif (1997) concluded that the hormonal contraceptive drug caused hyperactivity of lymphocyte function, this has been noticed by increase in the RNA and protein content in the cells. The administration of androgenic steroids in fish leads to a general increase in protein synthesis (Lone and Ince, 1983) and enhanced the proteolytic activity of the digestive tract (Lone and Matty, 1981). In mammals, anabolic steroids increase protein synthesis and activate the secretion (Michelsen et al., 1982). On the other hand, GH promotes both increased size of the cells and protein synthesis by the ribosomes, this is caused by a direct effect on ribosomes, stimulate the transcription of DNA causing formation of large quantities of RNA (Mountcastle, 1980). Sakai et al. (2004) stated that ghrelin was recently identified as an endogenous ligand for GH secretagogue receptor; in this study, they investigated the effects of ovariectomy on the numbers of ghrelin-immunopositive and -expressing cells, ghrelin mRNA levels and plasma ghrelin concentrations in female rats. Three days after ovariectomy, the number of ghrelin cells, plasma ghrelin level and the ghrelin mRNA level significantly increased. These responses were reversed by 17β-estradiol replacement. they also found that ghrelin-immunopositive cells express estrogen receptor. These results suggested that estrogen is involved in the regulation of ghrelin expression.
In the present study the amount of the mucus cells at the apical region of the gastric gland of the Nordette-treated animals was greatly decreased. The major functions of the gastric epithelium are the secretion of acid and digestive enzymes. It also secretes mucus to lubricate ingested food and to protect itself from the corrosive effects of the acid and enzymes. The mucosal surface of the stomach has a thick protective coating of mucus (Junqueira et al., 1995). In the present study, it has been noticed that Nordette cause damage to the mucus cell layer. Confirming the present study, Williams and Tumberg (1981) stated that a pH gradient across the mucus layer thereby established and acts as a protective barrier between the luminal acid and the gastric epithelium, therefore any damage to the mucus cells destruct the protective barrier. On the other hand, recent evidence suggests that tight junctions around surface and pit cells also form a part of barrier to acid. Stress and other psychosomatic factors or such substances as aspirin that cause gastric irritation can disrupt this epithelial layer and lead to ulceration (Junqueira et al., 1995).
In the present study, concerning the blood vessels, it appeared that an obvious adverse changes were observed in the blood vessels and blood capillaries including dilatation and congestion especially in the apical region of the gastric glands. Blood capillaries lined by enlarged endothelial cells containing enlarged nuclei with highly condensed heterochromatin. Endothelial cell cytoplasm contained lysosomes and ribosomes. Immunoblotting studies on the estrogens phosphorylate endothelial NOS (NOS III), suggesting that NOS III is the target for estrogen-induced NOS activity (Jaubert et al., 2007). Cells of the gastric epithelium are responsible for secretory and protective functions. To subserve these functions, an intimate relationship between these cells and the mucosal microcirculation might be expected. Physiological studies indeed show a direct relationship between acid secretion and gastric mucosal blood flow (Lanciault and Jacobson, 1976); abundant evidence implicates a distributed mucosal microcirculation in the pathogenesis of acute gastric ulceration (Moody, 1971). Progressing toward the gastric lumen there was a notable decrease in the amount of connective tissue between the basal lamina of the endothelium and that of the adjacent gastric gland cells. The capillaries situated immediately below the surface mucus cells were in close proximity to the basal laminae of these cells. Diffusion distances between capillaries and adjacent parietal cells of the gastric glands were usually even smaller (Gannon et al., 1982). In agreement with the above observations, the cytoplasm of oxyntic cells beside the blood capillaries contained many cytoplasmic vacuoles. This observation suggested that there is a disturbance in the diffusion of the small molecules between the capillary and the oxyntic cells.
The most pathological effects in the present study were restricted to the extensible decrease of the lymphoid cells in the submucosa and lamina propria. Although, there were many lymphoid cells lies in the submucosa and lamina propria of the stomach of the control animals the submucosa and lamina propria of the treated stomach were obviously decrease of lymphoid cells. Also, Yousif (1997) demonstrated that administration of a daily therapeutic dose of hormonal contraceptive drug for 30 days cause an obvious decrease of lymphocytes in the thymus and the spleen of the mice. Confirm to the above study, Jiang et al. (2006) showed that exposure to estrogens is associated with an increased risk of developing breast, cervical and liver cancer. Estrogens strongly induce the human granzyme B inhibitor, proteinase inhibitor 9 (PI-9). Because cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells use the granzyme pathway to induce apoptosis of target cells, Jiang et al. (2006) tested the ability of activated CTLs and the human NK cell line, YT cells, to lyse human liver cells. Estrogen induction of PI-9 protected the liver cells against CTL and NK cell-mediated, granzyme-dependent, apoptosis. Knockdown of PI-9 by RNA interference blocked the protective effect of estrogen. This study demonstrates that estrogens can act on target cells to control their destruction by immune system cells and shows that induction of PI-9 expression can inhibit both CTL and NK cell-mediated apoptosis. Estrogen induction of PI-9 may reduce the ability of cytolytic lymphocytes-mediated immune surveillance to destroy newly transformed cells, possibly providing a novel mechanism for an estrogen-mediated increase in tumor incidence (Jiang et al., 2006).
On the other hand, in the present study Nordette cause irregularity in cell boundaries of gastric gland cells. Recently, Li et al. (2006) stated that estrogen depletion decreased and treatment with 17β-estradiol increased deformability of cervical-vaginal epithelial cells. Estrogen also induced redistribution of nonmuscle myosin II-B (NMM-II-B); lesser interaction of NMM-II-B with actin; increased phosphorylation of NMM-II-B-heavy chains at threonine and serine residues and decreased filamentation of NMM-II-B in vitro. Augmented phosphorylation of NMM-II-B can block filamentation and induce disassociation of the myosin from the cortical actin and disruption of the actomyosin ring can increase cell deformability, we confirm these observations.
Ueyama et al. (2006) showed that gastic parietal cells produce and secrete a substantial amount of estrogen (E2) into portal vein, using biochemical and morphological step-up of E2 concentration in the portal vein, compared with that in the artery, which was not seen after gastrectomy and also an increase of arterial E2 concentration after portal-systemic shunting. This indicates that the stomach, but not other organs, is the main origin of portal venous E2. In the physiological tissue culture system, gastric mucosa produced testosterone and E2 from androstenetione. These data indicate the presence of aromatase in gastric mucosa. Because gastric mucosa contains a mixed population of cells. The histochemical data indicate that parietal cells in gastric mucosa have aromatase mRNA and its protein. Taken together, Ueyama et al. (2006) further showed that the signals for ERmRNA and protein were located in hepatocytes and mRNA levels for hepatic ER were regulated by E2 concentration in the blood flowing into the liver. Ueyama et al. (2006) proposed an expanded role of gastric parietal cells beyond the well-known exocrine function of secreting hydrochloric acid and intrinsic factor into gastric juice, that the gastric parietal cells serve an endocrine function, whereby estrogen is synthesized and secreted into the portal vein, the consequences of which may be of clinical significance in the case of portal-systemic shunting. Gastric parietal cells are unique, in that they secrete E2 and can produce its precursor, testosterone. Le Goascogne et al. (1995) reported that gastric parietal cells may use circulating progesterone derived from adrenal cortex and gonads as the first precursor for successive steps, or they can also use circulating testosterone preferentially as the substrate of aromatase. Gastric parietal cells are also endowed with a large number of mitochondria and an elaborate tubulovesicular system, similar to a smooth-surfaced endoplasmic reticulum. These structural characteristics seem to be common in adrenal cortical cells and testicular interstitial cells. A functional role for gastric estrogen has still to be elucidated. As one possibility, gastric estrogen may act as a local regulator of the gastro-hepatic axis, because ERα mRNA and immunoreactive protein were expressed in hepatocytes and mRNA levels of hepatic ERα were regulated by E2 concentration in the blood flowing into the liver. Estrogen might affect gastric motility (Bond et al., 1998), because a low level of ERα mRNA was expressed in the muscularis externa of stomach. Gastric estrogen might also act directly on parietal cells in an autocrine or intracrine fashion, because the protein and mRNA expression of ERα and ERβ was reported in gastric parietal cells (Campbell-Thompson et al., 2001).
By immunohistochemistry the expression of ER beta was studied by Matsuyama et al. (2002) in 29 stomach adenocarcinomas. In this study, the expression of ER beta in stomach adenocarcinomas has been investigated for the first time. All adenocarcinomas, demonstrated clear ER beta nucleus staining. Lymphocytes, venous endothelial cells, smooth muscle and non-cancerous stomach glands also showed strong ER beta staining, Equivalent ER beta protein levels in cancerous and non-cancerous tissues, which was consistent with the results of immunohistochemical staining. The results imply that the effects of estrogen in stomach cancer, as well as those in normal stomach, may be mediated by ER beta (Matsuyama et al., 2002). In the present study, the microscopic changes observed in Lymphocytes, endothelial cells of the capillaries and smooth muscle cells confirm the above observations.
CONCLUSION
From the present study, we can concluded that the contraceptive drug induced extensive light and electron microscopic changes and these changes may lead to disturbance in the stomach functions, including disturbance in the secretory function of the parietal cells, the interaction between the blood and oxyntic cells and the stomach mobility. The important characteristic features is the obvious decrease in the mucus cell layer in the treated stomach. On the other hand, the contraceptive drug obviously induce adverse effect in the immune system cells by decrease the amount of the lymphoid cells in the stomach. These changes, which noticed in gastric gland cells, lymphocytes, endothelial cells and smooth muscle fibers in the stomach may be mediated by ER. Contraceptive drug cause histolpathogical changes in the stomach and it may be toxic to the stomach when used for long period.
REFERENCES
- Ackerman, G.E., M.E. Smith, C.R. Mendelson, P.C. Macdonald and E.R. Simpson, 1981. Aromatization of androstenedione by human adipose tissue stromal cells in monolayer culture. J. Clin. Endocrinol. Metab., 53: 412-417.
CrossRef - Bond, E.F., M.M. Heitkemper and S.L. Bailey, 1998. Estrogen suppresses gastric motility response to thyrotropin-releasing hormone and stress in awake rats. Res. Nurs. Health, 21: 221-228.
PubMed - Brodie, A. and S. Inkster, 1993. Aromatase in the human testis. J. Steroid Biochem. Mol. Biol., 44: 549-555.
CrossRef - Campbell-Thompson, M., K.K. Reyher and L.B. Wilkinson, 2001. Immunolocalization of estrogen receptor and in gastric epithelium and enteric neurons. Endocrinology, 171: 65-73.
Direct Link - Farrell, G.C., A. Koltai and M. Murray, 1988. Source of raised serum estrogens in male rats with portal bypass. J. Clin. Invest., 81: 221-228.
CrossRef - Gambrell, Jr. R.D., 1992. Update on hormone replacement therapy. Am. Fam. Phys., 46: 87S-96S.
Direct Link - Gannon, B., J. Browning and P. O`Brien, 1982. The microvascular architecture of the glandular mucosa of rat stomach. J. Anat., 135: 667-683.
Direct Link - Gorodeski, G.I., 2007. Estrogen decrease in tight junctional resistance involves matrix-metalloproteinase-7-mediated remodeling of occludin. Endocrinology, 148: 218-231.
Direct Link - Harada, N., H. Sasano, H. Murakami, T.Ohkuma, H. Nagura and Y. Takagi 1999. Localized expression of aromatase in human vascular tissues. Circ. Res., 84: 1285-1291.
PubMed - Jaubert, A., N. Mehebik-Mojaat, D. Lacasa, S. Dominique, G. Yves and R. Catherine, 2007. Nongenomic estrogen effects on nitric oxide synthase activity in rat adipocytes. Endocrinology, 148: 2444-2452.
Direct Link - Jiang, X., B.A. Orr, D. Kranz and D.J. Shapiro, 2006. Estrogen induction of the granzyme B inhibitor, proteinase inhibitor 9, protects cells against apoptosis mediated by cytotoxic T lymphocytes and natural killer cells. Endocrinology, 147: 1419-1426.
Direct Link - Le Goascogne, C., N. Sananes, B. Eychenne, M. Gouezou, E.E. Baulieu and P. Robel, 1995. Androgen biosynthesis in the stomach: Expression of cytochrome P450 17 alpha-hydroxylase/17,20-lyase messenger ribonucleic acid and protein and metabolism of pregnenolone and progesterone by parietal cells of the rat gastric mucosa. Endocrinology, 136: 1744-1752.
CrossRefDirect Link - Lephart, E.D., 1996. A review of brain aromatase cytochrome P450. Brain Res. Rev., 22: 1-26.
CrossRef - Leshin, M., J. Baron, F.W. George and J.D. Wilson, 1981. Increased estrogen formation and aromatase activity in fibroblasts cultured from the skin of chickens with the Henny-feathering trait. J. Biol. Chem., 256: 4341-4344.
Direct Link - Li, X., L. Zhou and G.I. Gorodeski, 2006. Estrogen regulates epithelial cell deformability by modulation of cortical actomyosin through phosphorylation of nonmuscle myosin heavy-chain II-B filaments. Endocrinology, 147: 5236-5248.
Direct Link - Lone, K.P. and A.J. Matty, 1981. The effect of feeding androgenic hormones on the proteolytic activity of the alimentary canal of carp Cyprinus carpio L. J. Fish. Biol., 18: 353-358.
CrossRefDirect Link - Lone, K. and B. Ince, 1983. Cellular growth response of rainbow trout (Salmo gairdneri) fed different levels dietary protein and anabolic steroid hormone ethylestrenol. Gen. Comp. Endocrinol., 49: 32-49.
CrossRef - Matsuyama, S., Y. Ohkura, H. Eguchi, K. Yasuhito and A. Kiwamu et al., 2002. Estrogen receptor beta is expressed in human stomach adenocarcinoma. J. Cancer Res. Clin. Oncol., 128: 319-324.
Direct Link - Michelsen, C., J. Askanazi and J. Kinney, 1982. Effect of an anabolic steroid on nitrogen balance and amino acid patterns after total replacement. J. Trauma, 22: 410-413.
PubMed - Purohit, A., A.M. Flanagan and M.J. Reed, 1992. Estrogen synthesis by osteoblast cell lines. Endocrinology, 131: 2027-2029.
CrossRef - Sakai, T., K. Inoue, M. Yamazaki, I. Kinji and S. Takafumi, 2004. Estrogen modulates ghrelin expression in the female rat stomach. Peptides, 25: 289-297.
Direct Link - Samy, T.S., M.W. Knöferl, R. Zheng, G. Martin Schwacha, I. Kirby Bland and H. Irshad Chaudry, 2001. Divergent immune responses in male and female mice after trauma-hemorrhage: Dimorphic alterations in T lymphocyte steroidogenic enzyme activities. Endocrinology, 142: 3519-3529.
Direct Link - Simpson, E., G. Rubin, C. Clyne, K. Robertson, L. O'Donnell, S. Davis and M. Jones, 1999. Local estrogen biosynthesis in males and females. Endocrinol. Relat. Cancer, 6: 131-137.
CrossRefDirect Link - Stallard, S., J. Litherland, C. Cordiner, M. Hilary Dobson and W. David George et al., 2000. Effect of hormone replacement therapy on the pathological stage of breast cancer: Population based, cross sectional study. Br. Med. J., 320: 348-349.
Direct Link - Ueyama, T., M. Shirasawa, N. Numazawa, Y. Keiko, S. Momoko, I. Takao and T. Yoshihiro, 2006. Gastric parietal cells: Potent endocrine role in secreting estrogen as a possible regulator of gastro-hepatic axis. Endocrinology, 143: 3162-3170.
Direct Link - Vandenberg, L.N., M.V. Maffini and P.R. Wadia, 2007. Exposure to environmentally relevant doses of the xenoestrogen Bisphenol-A alters development of the fetal mouse mammary gland. Endocrinology, 148: 116-127.
CrossRef - Williams, S.E. and L.A. Tumberg, 1981. Demonstration of a pH gradient across mucus adherent to rabbit gastric mucosa: Evidence for a mucus bicarbonate' barrier. Gut, 22: 94-96.
Direct Link