
E. Farshadfar
College of Agriculture, Razi University, Kermanshah, Iran
H. Ghasempour
College of Science, Razi University, Kermanshah, Iran
H. Vaezi
College of Agriculture, Razi University, Kermanshah, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 1 | Page No.: 118-122







ABSTRACT
In order to evaluate molecular indices of drought tolerance and selection of drought tolerance genotypes in bread wheat a randomized complete block design was carried out with three replications in the field of research station of Sararood, Kermanshah, Iran. Positive significant correlation coefficient was observed between grain yield and proline, soluble sugar and total protein under rainfed condition. Based on the grain yield, proline content, soluble sugar and total protein the genotypes were classified into four clusters using cluster analysis and UPGMA method. Genotypes numbers 4, 9, 10, 11, 18 and 19 in the first clusters indicated the highest performance.
PDF Abstract XML References Citation
How to cite this article
E. Farshadfar, H. Ghasempour and H. Vaezi, 2008. Molecular Aspects of Drought Tolerance in Bread Wheat (T. aestivum). Pakistan Journal of Biological Sciences, 11: 118-122.
DOI: 10.3923/pjbs.2008.118.122
URL: https://scialert.net/abstract/?doi=pjbs.2008.118.122
DOI: 10.3923/pjbs.2008.118.122
URL: https://scialert.net/abstract/?doi=pjbs.2008.118.122
INTRODUCTION
Water deficit is the most important factor in yield reduction in the semiarid regions (Kristin et al., 1997; Kirigwi et al., 2004; Farshadfar and Sutka, 2003).Positive correlation with relative yield performance of genotypes in rainfed condition is a start point for identification of characters related to drought tolerance (Farshadfar and Sutka, 2003; Ghasempour and Kianian, 2007).Drought tolerance is not easily quantifiable plant attribute, rather it is the outcome of interaction among complex morphological, physiological and molecular character associated with low molecular weight biochemical materials acting as osmoticum, these include proline, betaine, choline, betain aldeid, 3 etil butarat and dietil glycerin (Khazaei, 2001; Ghasempour et al., 2007; Ildiko and Galiba, 2000).Under drought stress, high molecular weight (>100kd) soluble protein will decrease in wheat leaves, while low molecular weight will increase (Kochaki, 1997; Sujin and RayWu, 2004). Drought stress decompose starch and fade it from the plant. Reduction of starch is the result of amylase activity that increase soluble sugar (Vaezi, 2005; Ghasempour et al., 1998). Genotypes with high soluble sugar show drought resistance. Decreases phospholipids in the cell membrane (Zarei, 2006) suggesting biochemical attributes such as: free proline content, soluble sugar, total protein and chlorophyll stability can be used as drought tolerance indicators for selecting drought resistant genotypes (Khazaei, 2001; Sujin and RayWu, 2004).The objectives of the present investigation were to screening molecular indicators of drought tolerance and selection of drought tolerant genotypes.
MATERIALS AND METHODS
In order to screen drought tolerant genotypes twenty fall bread wheat cultivars were incubated at 4°C for one week for vernalization and evaluated in a randomized complete block design with three replications under drought and irrigated conditions in the field. Each plot consisted of 3 rows with row to row distance of 50 cm.
In the rainfed condition besides Grain Yield (GY), from each experimental unit 5 plants were randomly selected and the following metabolic traits were measured from the flag leaves:
Soluble sugar extraction: Soluble sugar content was determined by the modified phenol sulphuric acid method (Dubois et al., 1956; Kennedy, 1987). Data were measured at 485 nm by Bausch and Lomb spectrophotometer 70. A standard curve; 0, 5, 10, 15, 20, 25, 30 and 40 mg of glucose were prepared. Glucose content of treated extracts was calculated using the standard curve and recorded.
Assessment of proline in leaves: Proline of leaves was determined by Bates et al. (1973) method. Data were measured at 520 nm by Bausch and Lomb spectrophotometer 70. A standard curve; 1.9, 7.8, 15.62, 31.25 and 125 μg of proline were prepared. Proline content of treated extracts was calculated using the standard curve and recorded.
Measuring the total protein (T. protein): In order to quantify the total protein of leaves in each sample, a 0.05 g of dry weight of leaves was evaluated by Lowry et al. (1951) method. The total protein was determined with Folin reagent and the color compared with Bovine Serum Albumin (BSA), serving as the standard for determining protein content, read at A660 (OD) and recorded.
Protein SDS-PAGE and gel electrophoresis analysis: Peterson (1977) method was used to determine the protein concentration of fresh leaves of treated plants.
For one-dimensional SDS-PAGE the supernatant of samples was diluted with UKS-buffer (9.5 Urea, 5 mM K2CO3, 1.25% (w/v) SDS) (1:1).
For each well 20 μL was applied. A Hoeffer SE 600 vertical unit was employed and coomassie blue used for staining.
In order to analyze molecular weight and mobility of protein bands the UV. Doc program was used and molecular weight of marker bands entered to the program. The program gives the molecular weight of the bands on the stained gel.
Chlorophyll content: Using Ashraf et al. (1994) method spectrophotometer with 663 and 645 nm and the following formulas, chlorophyll content was determined:
Chlorophyll a (Ch.a) (mg mL-1): 0.0202a645+0.008a663
Chlorophyll b (Ch.b)(mg mL-1): 0.0127a663- 0.00269a645
Total chlorophyll (T.Ch.) (mg mL-1): 0.0229a645- 0.00468a663
Statistical analysis: Analysis of variance, mean comparison, correlation and cluster analysis were done with MSTAT-C and SPSS statistical softwares.
RESULTS AND DISCUSSION
The results of analysis of variance under water stress condition (Table 1) revealed high significant differences between genotypes for proline content, soluble protein, chlorophyll a, b and total chlorophyll and grain yield indicating the presence of genetic variation and possibility of selection for drought tolerant genotypes.
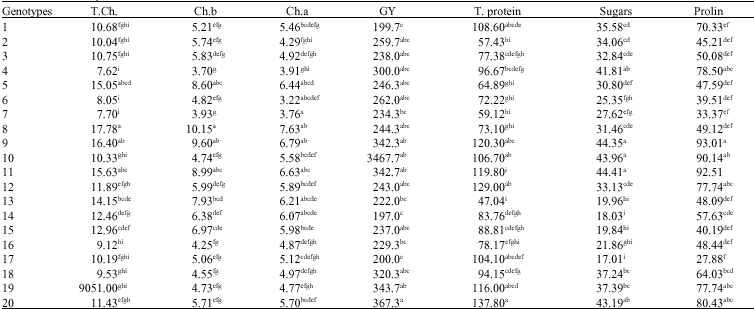
Mean comparison (Table 2) for total chlorophyll ranged from 7.62 mg mL-1 for genotype 4 to 17.78 mg mL-1 for genotype 8, chlorophyll a ranged from 3.22 for genotype 6 to 7.63 for genotype 8 and chlorophyll b ranged from 3.70 for cultivar 4 to 10.15 for cultivar 8. Maximum total protein (137.8) and grain yield (367.3 g) was related to genotype 20. Genotypes number 9 and 11 exhibited maximum amount of proline (93.01) and soluble sugar (44.41), respectively.
Proline content and soluble sugar were larger during the grain filling period than preanthesis, hence the best
| Table 1: | Analysis of variance for various characters investigated under water stress condition |
 | |
| ns: non significant; **: Significant at 0.01 probability level; +: Error = 38 | |
| Table 2: | Mean comparisons of characters studied under water stress condition + |
 | |
| +: Genotypes with common letter(s) have no significant differences | |
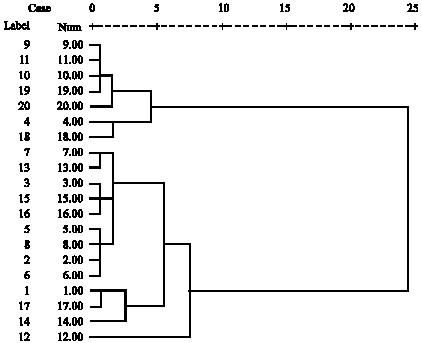 | |
| Fig. 1: | Cluster analysis based on the characters with positive correlation with grain yield |
| Table 3: | Correlation coefficients between characters studied |
 | |
| *, **: Significant at 5 and 1% level of probability, respectively | |
stage for selection of drought tolerant genotypes will be postanthesis and grain filling period (Hien et al., 2003).
With regard to this fact cultivars number 9, 10, 11, 12 and 20 have the highest amount of proline content, soluble sugar and total protein.
As chlorophyll has an important role in photosynthesis rate, therefore, genotypes with high amount of photosynthesis in grain filling period will have higher grain yield and resistant to water stress. With this regard and what mentioned before, genotype number 11 will be outstanding regarding grain yield and all metabolite and physiological traits.
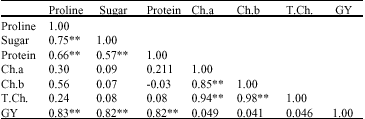
Correlation analysis: One criterion for a character to be an index of drought tolerance is having positive significant correlation coefficient with grain yield under water stress (Sujin and Ray Wu, 2004), as proline, soluble sugar and protein showed high significant correlation coefficients with grain yield under drought condition (Table 3), therefore they can be considered as drought tolerance indicators (Ghasempour et al., 2001).
Soluble sugar revealed significant positive correlation coefficient with proline (r = 0.750**) and protein (r = 0.66**). Which is in accordance with Siegien and Leszczynska (2004) and Zarei (2006). Proline and soluble protein also exhibit positive high significant correlation coefficient (r = 0.663**).
It can be concluded that effect of water stress on genotypes with higher soluble protein causes protein degradation and release more free proline (Vaezi, 2005). Chlorophyll a and b and total chlorophyll did not show any significant correlation with grain yield and other characters, hence according to the results of this investigation they can not be considered as drought tolerance indicators.
Based on the characters having significant variation and significant positive correlation with grain yield (proline, soluble sugar and protein) cluster analysis using UPMGA showed four clusters so that high yielding and drought tolerant genotypes 4, 9, 10, 11, 18, 19, 20 with regard to biochemical traits were grouped in cluster one (Fig. 1). Genotypes number 1, 14, 17 and 12 were grouped in clusters 3 and 4. As genotype 1, 12, 14 and 17 (cluster 3 and 4) and genotypes 4, 9, 10, 11, 18, 19 and 20 (cluster one) have the most genetic distance, hence their crossing will contribute to heterosis and transgressive segregating in the segregation populations (Altinkut et al., 2001; Vaezi, 2005).
Evaluation of width and number of protein bands in SDS-PAGE indicates that after stress 12 protein bands appeared.
With regard to this fact that each protein band is an indicator of one character, therefore different between protein bands exhibited genetic variation between wheat genotypes.
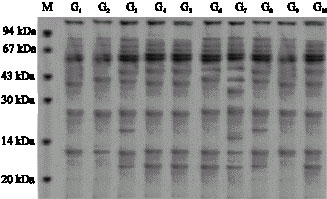 | |
| Fig. 2: | Protein profile (1-10) genotypes (before stress) |
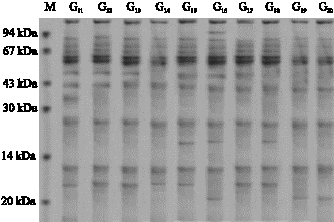 | |
| Fig 3: | Protein profile (11-20) genotypes (before stress) |
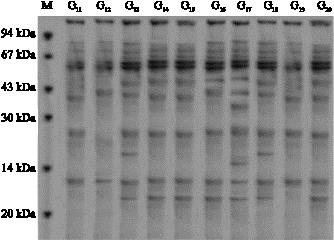 | |
| Fig. 4: | Protein profile (1-10) genotypes (next stress) |
Figure 2-5 also revealed that in some genotypes some new bands appeared and some disappeared. For example in genotype 1 the first band at the end appeared after water stress, while in genotype 2 the first band at the end disappeared. In genotype 3 a new band appeared, in genotype 7 third band disappeared, in genotype 19 and 1 appeared. In genotypes No. 1, 2, 3 an 4 the intensity of
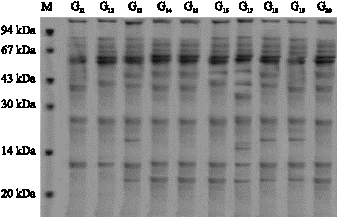
band No. 2 and in genotypes 13, 14, 15 and 16 the intensity of band No. 3 increased while in genotype 12 the intensity of band No. 3 decreased. From the above mentioned results it is concluded that the increase of the intensity of bands is the results of the increase of proteins with low molecular weight (Zimmermann, 1998) reported that water stress in wheat leaves causes soluble proteins with low molecular weight increase while, soluble proteins with high molecular weight decrease (Vaezi, 2005; Ghasempour et al., 2007).
REFERENCES
- Altinkut, A., K. Kazan, Z. Ipekci and N. Gozukirmizi, 2001. Tolerance to paraquat is correlated with the traits associated with water stress tolerance in segregating F2 populations of barley and wheat. Euphytica, 121: 81-86.
CrossRefDirect Link - Ashraf, M.Y., A.R. Azemi, A.H. Khan and S.A. Ala, 1994. Effect of water stress and total phenols, peroxidase activity and chlorophyll content in wheat. Acta Physiol. Plant., 16: 185-191.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Farshadfar, E. and J. Sutka, 2003. Multivariate analysis of drought tolerance in wheat substitution line. Cereal. Res. Commun., 31: 33-40.
Direct Link - Ghasempour, H.R., D.F. Gaff, R.P.W. Williams and R.D. Gianello, 1998. Contents of sugars in leaves of drying desiccation tolerant flowering plants, particularly grasses. Plant Growth Reg., 24: 185-191.
CrossRef - Ghasempour, H.R., E.M. Anderson and F. Gaff Donald, 2001. Effects of growth substances on the protoplasmic drought toleranceof leaf cells of the resurrection grass, Sporobolus stapfianus. Aust. J. Plant Physiol., 28: 1115-1120.
Direct Link - Ghasempour, H.R. and J. Kianian, 2007. The study of desiccation-tolerance in drying leaves of the desiccation- tolerance grass Sporobolus elongatus and the desiccation-sensitive grass Sporobolus pyramidalis. Pak. J. Biol. Sci., 10: 797-801.
CrossRefPubMedDirect Link - Hien, D.T., M. Jacobs, G. Angenon, C. Hermans, T.T. Thu, L. van Son and N.H. Roosens, 2003. Proline accumulation and Δ1-pyrroline-5-carboxylate synthetase gene properties in three rice cultivars differing in salinity and drought tolerance. Plant Sci., 165: 1059-1068.
Direct Link - Ildiko, K. and G. Galiba, 2000. Osmotic and salt stress- induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci., 40: 482-487.
Direct Link - Kirigwi, F.M., M. van Ginkel, R. Trethowan, R.G. Sears, S. Rajaram and G.M. Paulsen, 2004. Evaluation of selection strategies for wheat adaptation across water regimes. Euphytica, 135: 361-371.
CrossRefDirect Link - Kristin, A.S., R.R. Serna, F.I. Perez, B.C. Enriquez and J.A.A. Gallegos et al., 1997. Improving common bean performance under drought stress. Crop Sci., 37: 51-60.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Peterson, G.L., 1977. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem., 83: 346-356.
CrossRefPubMedDirect Link - Siegien, N. and R.B. Leszczynska, 2004. Effects of drying rate on dehydration sensitivity of excised wheat seedling shoots as related to sucrose metabolism and antioxidant enzyme activities. Plant Sci., 167: 879-888.
Direct Link - Su, J. and R. Wu, 2004. Stress-inducible synthesis of proline in transgenic rice confers faster growth under stress conditions than that with constitutive synthesis. Plant Sci., 166: 941-948.
CrossRefDirect Link - Zimmermann, M.H., 1998. Hydraulic architecture of some diffuse porpous trees. Can. J. Bor., 56: 2286-2295.
CrossRefDirect Link - Altinkut, A., K. Kazan, Z. Ipekci and N. Gozukirmizi, 2001. Tolerance to paraquat is correlated with the traits associated with water stress tolerance in segregating F2 populations of barley and wheat. Euphytica, 121: 81-86.
CrossRefDirect Link - Ashraf, M.Y., A.R. Azemi, A.H. Khan and S.A. Ala, 1994. Effect of water stress and total phenols, peroxidase activity and chlorophyll content in wheat. Acta Physiol. Plant., 16: 185-191.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Farshadfar, E. and J. Sutka, 2003. Multivariate analysis of drought tolerance in wheat substitution line. Cereal. Res. Commun., 31: 33-40.
Direct Link - Ghasempour, H.R., D.F. Gaff, R.P.W. Williams and R.D. Gianello, 1998. Contents of sugars in leaves of drying desiccation tolerant flowering plants, particularly grasses. Plant Growth Reg., 24: 185-191.
CrossRef - Ghasempour, H.R., E.M. Anderson and F. Gaff Donald, 2001. Effects of growth substances on the protoplasmic drought toleranceof leaf cells of the resurrection grass, Sporobolus stapfianus. Aust. J. Plant Physiol., 28: 1115-1120.
Direct Link - Ghasempour, H.R. and J. Kianian, 2007. The study of desiccation-tolerance in drying leaves of the desiccation- tolerance grass Sporobolus elongatus and the desiccation-sensitive grass Sporobolus pyramidalis. Pak. J. Biol. Sci., 10: 797-801.
CrossRefPubMedDirect Link - Ghasempour, H.R., A.A. Hojatjalali and A.R. Rangin, 2007. Physiological changes, proline, total protein, protein analysis and potassium of the sugar beet plants in Response to Beet cyst Nematodes, Heterodera schachtii. Int. J. Bot., 3: 91-96.
Direct Link - Hien, D.T., M. Jacobs, G. Angenon, C. Hermans, T.T. Thu, L. van Son and N.H. Roosens, 2003. Proline accumulation and Δ1-pyrroline-5-carboxylate synthetase gene properties in three rice cultivars differing in salinity and drought tolerance. Plant Sci., 165: 1059-1068.
Direct Link - Ildiko, K. and G. Galiba, 2000. Osmotic and salt stress- induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci., 40: 482-487.
Direct Link - Kirigwi, F.M., M. van Ginkel, R. Trethowan, R.G. Sears, S. Rajaram and G.M. Paulsen, 2004. Evaluation of selection strategies for wheat adaptation across water regimes. Euphytica, 135: 361-371.
CrossRefDirect Link - Kristin, A.S., R.R. Serna, F.I. Perez, B.C. Enriquez and J.A.A. Gallegos et al., 1997. Improving common bean performance under drought stress. Crop Sci., 37: 51-60.
Direct Link - Peterson, G.L., 1977. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal. Biochem., 83: 346-356.
CrossRefPubMedDirect Link - Siegien, N. and R.B. Leszczynska, 2004. Effects of drying rate on dehydration sensitivity of excised wheat seedling shoots as related to sucrose metabolism and antioxidant enzyme activities. Plant Sci., 167: 879-888.
Direct Link - Su, J. and R. Wu, 2004. Stress-inducible synthesis of proline in transgenic rice confers faster growth under stress conditions than that with constitutive synthesis. Plant Sci., 166: 941-948.
CrossRefDirect Link - Zimmermann, M.H., 1998. Hydraulic architecture of some diffuse porpous trees. Can. J. Bor., 56: 2286-2295.
CrossRefDirect Link