
Fatemeh Bena Kashani
Department of Weed Research, Plant Pest and Disease Research Institute, P.O. Box 19395-1454, Tehran, Iran
Hasan Mohammad Alizadeh
Department of Agronomy, Agriculture and Natural Sciences Campus, University of Tehran, Karaj, Iran
Eskandar Zand
Department of Weed Research, Plant Pest and Disease Research Institute, P.O. Box 19395-1454, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 1 | Page No.: 72-77







ABSTRACT
Greenhouse and laboratory experiments were performed to evaluate the resistant of wild oat Avena luduviciana Durieu. populations to fenoxaprop-p-ethyl. Populations of A. ludoviciana were collected from different locations in Iran, showed indications of resistance to this herbicide. Whole plant assay experiments included screening tests and dose response experiments whereas; seed bioassay experiment consisted of ID50 determination and dose response experiments. Whole plant assay experiments were conducted as a randomized complete block design in four replications. The treatments were wild oat populations included FR1, FR2, FR3, FR4 (collected from Fars province), MR1, MR2, MR3 (collected from Markazi province), KS, KR1, KR2, KR3 (collected from Khuzestan province) and S (collected from location which had never been treated previously with any graminicide). Seed bioassay experiments were conducted using a randomized design with 4 replications. On the whole plant basis, resistance was found in, KR1, KR2, KR3 and FR4 and based on a seed bioassay, these populations were also resistant to fenoxaprop-p-ethyl. Resistance ratios (R/S) of resistant populations were different. Present findings also revealed that the seed bioassay could be used as a simple, comparatively rapid, inexpensive and accurate method for identifying wild oat populations resistant to Acetyl CoA carboxylase (ACCase) inhibitors.
PDF Abstract XML References Citation
How to cite this article
Fatemeh Bena Kashani, Hasan Mohammad Alizadeh and Eskandar Zand, 2007. Investigating the Resistance of Wild Oat (Avena ludoviciana Durieu.) to Fenoxaprop-p-ethyl by Whole Plant Bioassay and Seed Bioassay. Pakistan Journal of Biological Sciences, 10: 72-77.
DOI: 10.3923/pjbs.2007.72.77
URL: https://scialert.net/abstract/?doi=pjbs.2007.72.77
DOI: 10.3923/pjbs.2007.72.77
URL: https://scialert.net/abstract/?doi=pjbs.2007.72.77
INTRODUCTION
Since the introduction of the Aryloxyphen- Oxypropionate (APP) and Cychlohexanedione (CHD) herbicides of ACCase inhibitors in 1977, farmers have increasingly become reliant on these herbicides for grass weed control II. The increase in the use of ACCase inhibitors led to a parallel increase in the evolution of resistant populations to these herbicides (Rubin, 1996). To date, many grass weed species Lolium rigidum Gaudin., Phalaris minor Retz., Alopecurus myosuroides Hudson. and Avena fatua L. have been reported to be resistant to ACCase inhibiting herbicides. The resistance of wild oat to ACCase inhibitors was first reported in the Canadian prairies in 1990, 14 years after the registration of diclofop-methyl and 7 years after that of sethoxydim (Heap et al., 1993). Since then, resistance of wild oat to ACCase herbicides has been reported in many countries worldwide including, Canada, Australia, France, South Africa, United State and Chile (Heap, 2006).
Several methods have been developed to detect weed resistance to herbicides. They include whole plant assay, petri dish assay (Beckie et al., 1990), chlorophyll fluorescence, leaf disc flotation and enzyme sensitivity assays (Moss, 1995). The most widely used test for identifying herbicide resistance is whole plant assay (Moss, 1999). The determination method should be rapid, accurate, cheap and rapidly available and should provide a reliable indication of likely effect of resistance on herbicide performance in the field (Do-Soon et al., 2000). Seed bioassay is comparatively quick, inexpensive and is particularly useful for routine screening of large numbers of suspected resistant populations (Murray et al., 1996). Petri dish bioassays, which generally involve either shoot length or root length as growth parameters to discriminate between resistant and susceptible biotypes exposed to herbicide solutions, have already been developed to screen resistance within populations. Such bioassay has been successfully used for documenting green foxtail (Setaria viridis (L.) P. Beauv.) resistance to trifluralin (Beckie et al., 1990), wild oat (Avena fatua L.) resistance to the aryloxyphenoxypropionate and cychlohexanedione herbicides (Murray et al., 1996) and detection of propanil and fenoxaprop resistance in Echinochloa colonum (L.) Link. (Do-Soon et al., 2000).
In Iran, APP herbicides have continuously been used for selective control of wild oat and other grass weeds since 1980 (Zand and Baghestani, 2002). Recently, unsatisfactory control of wild oat (A. ludoviciana) using these herbicides has been reported from some wheat growing areas including Khuzestan, Fars and Markazi provinces, where wheat is grown. Poor control of this weed could not be attributed to improper application of these herbicides, but it may be due to evolution of herbicide resistance in A. ludoviciana populations at these locations. The objective of this study were (1) to determine whether A. ludoviciana populations in Iran have become resistant to fenoxaprop-p-ethyl and (2) to compare the efficiency of the whole plant assay with the seed bioassay for identifying herbicide resistance in weed populations.
MATERIALS AND METHODS
Plant material: Ten suspected resistant A. ludoviciana populations were collected in 2001 from wheat fields in Fars (FR1, FR2, FR3, FR4), Markazi (MR1, MR2, MR3) and Khuzestan (KR1, KR2, KR3) provinces. Seeds of the suspected resistant populations were collected from many plants that survived an annual treatment with aryloxyphenoxypropionate herbicides that had been used for at least 4-5 successive years. A susceptible (S) population was also collected from location which had never been treated previously with any graminicide (Tal et al., 2000). Populations were coded based on the province and susceptibility or suspicious to resistance (for example; KR1: suspicious to resistance population that was collected from Khuzestan province).
The present study consisted of two separate experiments, whole plant assay and seed bioassay experiments. Whole plant assay consisted of screening for resistance with fenoxaprop-p-ethyl and dose response experiments. Seed bioassay experiment included ID50 determination and dose response experiments. Both experiments were conducted at greenhouse facilities and laboratory of Plant Pest and Disease Research Institute, Tehran. It should be noted that all experiments were repeated twice.
Whole plant assay
Screening test: The experiment was conducted in a randomized complete block design with four replications. An individual pot containing 10 seeds was considered a treatment unit. Before planting and in order to break the seed dormancy, A. ludoviciana seeds were dehulled by hand and germinated on filter paper moistened with 8 mL distilled water in 9 cm plastic Petri plates. Plates were transferred to a refrigerator at +5°C in the dark for 24 h and then placed in a germinator at +20/10°C with a 16/8 h and darkness to germinate the seeds. Ten seeds of wild oat were planted at a depth of 1 cm in 12 cm diameter pots filled with a loam/sand/peat mixture in a 1:1:1 ratio. Pots were transferred to a greenhouse at 25/18°C day/night temperature regime. Pots were watered daily to field capacity.
Fenoxaprop-p-ethyl at 75 g ai ha-1 was applied on wild oat at the two- to three-leaf stage. Herbicide were sprayed in a cabinet sprayer equipped with a flat-fan nozzle calibrated to deliver 200 L ha-1 of spray solution at a pressure of 2 bar. Visual percent wild oat control was rated 28 Day After Herbicide Application (DAHA) using EWRC rating system (Sandral et al., 1997). Four weeks after treatment, number of survived plants in each pot was counted, then the plants were harvested and oven dried at 75°C for 48 h and weighted. Percent wild oat biomass was calculated by dividing plant biomass in the untreated pot by plant biomass in the untreated pot and multiplying by 100. Those populations that were distinguished as resistant were studied further in a dose-response experiment to determine the level of resistance to fenoxaprop-p-ethyl.
Dose-response experiment: Dose response experiment was conducted using 12*10 cm deep pots in a randomized complete block design with four replications. Preparation of planting material and seed germination condition were similar to screening test. The A. ludoviciana populations that were selected in the previous experiment were tested at a range of fenoxaprop-p-ethyl doses. The applied fenoxaprop-p-ethyl doses were 0, 7.5, 18.75, 37.5, 75, 150, 300, 450, 600 and 1200 g ai ha-1, that covered rang of 0.1 to 16 recommended doses.
Seed bioassay
Discriminating dose experiment: The objective of this experiment was to determine herbicide dose at which 50% coleoptile’s length of susceptible population reduces (ID50), as a discriminating dose between resistant and susceptible populations. The experiment was performed as a completely randomized design with four replications. 10 imbibed seeds of S population (S) were placed over a filter paper in each Petri dish. Eight milliliter aqueous emulsion of commercially formulated fenoxaprop-p-ethyl was applied at a range of doses to sheet of filter paper lining the bottoms of petri plates. Then applied doses used were 0, 0.1, 0.2, 0.4, 0.8 and 1 mg L-1. Petri plates were kept for 48 h in the dark in a germination cabinet with a day/night temperature regime at 20/10°C, respectively. The coleoptile’s lengths were measured after 7 days. After determining the ID50 of susceptible population, discriminating dose was applied to all populations.
Dose-response experiments: Dose response experiment was arranged as a completely randomized design with four replications. Seeds preparation and germination were the same as described in discriminating dose experiment. Fenoxaprop-p-ethyl was applied at doses of 0, 0.1, 0.2, 0.4, 0.8 and 1 mg L-1.
All data were subjected to analysis of variance (ANOVA) using SAS software (SAS Institute, 1996). The assumptions of the variance analysis were tested by insuring that the residuals were random, homogeneous with a normal distribution about a mean of zero. If the assumptions of variance were not adequately met, data were subjected to an arcsin square root transformation (for data calculated as percent of the check treatment) or square root transformation (for visual rating scores). A nonlinear regression equation (Brain and Cousens, 1989) was fitted to dose-response data and used to describe the response of the populations to fenoxaprop-p-ethyl:
Y = k/(1+ ebg xb)+d |
Where Y is dependent variable, x is the herbicide dose, e is the base of natural logarithm, k is the difference between the upper and lower asymptotes, k+d is the upper asymptote, d is the lower asymptote and b and g determine the shape of the curve. Regression equations were used for calculating herbicide application rates required to inhibit growth, surviving plant and to inhibit coleoptile’s length by 50% (ID50). Resistance ratios (R/S) were then calculated by dividing the ID50 of the resistant populations by the susceptible population.
RESULTS AND DISCUSSION
Whole plant assay
Screening test: At 28 DAHA, A. ludoviciana biomass, survival and visual injury were significantly different among the populations when treated by fenoxaprop-p-ethyl (Table 1). KR1, KR2 and KR3 showed the least biomass reduction and the highest plant survival, while other populations were satisfactorily controlled by fenoxaprop-p-ethyl. These results are in agreement with the results of visual injury. According to Beckie et al. (2000), a population would be considered as resistant if show survival at least 50% and be able to keep its biomass at least 80% the untreated check.
| Table 1: | Wild oat shoot biomass and survived plant and visual percent weed control, 4 weeks after fenoxaprop-p-ethyl application at whole plant assay experiment and coleoptile's length 7 day after herbicide application at seed bioassay |
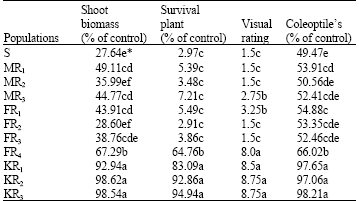 | |
* In each column, means with same letter(s) do not differ at 0.05 probability level according to Duncan multiple range test | |
However, when biomass reduces to 50% the untreated check, the population could be considered as possibly resistant. As a result, KR1, KR2 and KR3 were resistant to fenoxaprop-p-ethyl while FR4 was grouped as possibly resistant.
As observed, did not confirm our initial assumption about suspected resistance of Markazi and Fars populations. This indicates that control of A. ludoviciana at these locations would be attributed to other reasons like improper application, timing or method.
In this study, 75 g ai ha-1 fenoxaprop-p-ethyl was considered as discriminating dose because it is the recommended dose fenoxaprop-p-ethyl for wild oat control and farmers almost use this rate for control of wild oat, especially in wheat fields.
Dose-response experiments: As observed in the screening test, KR1, KR2, KR3 were chosen as resistant and FR4 was chosen as suspected resistant population. Population S also considered as the susceptible population. In dose-response experiment the relationship between shoot biomass and survival in these populations and fenoxaprop-p-ethyl doses were described by a sigmoidal model (Fig. 1 and 2). The dose response experiment showed that differences in shoot biomass and survival between the resistant and susceptible populations over all the range doses (Fig. 1 and 2). Among the populations, KR3 was the highest resistant population. At the highest dose of fenoxaprop-p-ethyl (1200 g ai ha-1) that was applied in this experiment, shoot biomass of KR3 population was 54.77% of control. Shoot growth of S population was also inhibited (48.26% of control) at half recommended dose (37.5 g ai ha-1) (Fig. 1). R/S ratios indicated that although all populations were resistant to fenoxaprop-p-ethyl, but there were clear differences in the level of resistance (Table 2 and 3).
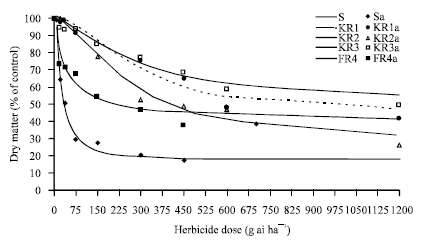 | |
| Fig. 1: | Effect of different fenoxaprop-p-ethyl concentrations on shoot biomass of susceptible (S) and resistant (KR1, KR2, KR3, FR4) populations of wild oat, as a percentage of untreated control. Symbols and lines represent actual and estimated responses, respectively |
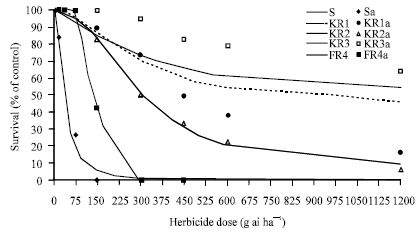 | |
| Fig. 2: | Effect of different fenoxaprop-p-ethyl concentrations on survival of susceptible (S) and resistant (KR1, KR2, KR3, FR4) populations, as a percentage of untreated controls. Symbols and lines represent actual and estimated response, respectively |
| Table 2: | Parameter estimates of the shoot biomass of susceptible and resistant populations as a percentage of untreated control, 4 weeks after fenoxaprop-p-ethyl application. Data were fitted according to the non-linear regression model: Y = k/(1ebg xb)+d* |
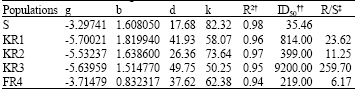 | |
*Y: dependent variable, x: The herbicide dose, e: The base of natural logarithm, k: The difference between the upper and lower asymptotes, k+d: The upper asymptote, d: The lower asymptote, b and g: The shape of the curve, +Herbicide application rates required to inhibite growth by 50%, +Dividing ID50 of the resistant populations by the susceptible population | |
KR3 largely differed from other populations in this respect since its ID50 was more than 259 times population S (Table 2). KR3 level of resistance to fenoxaprop-p-ethyl is comparable with very high levels of resistance to ACCase inhibitors in some populations of rigid rye grass (Lolium rigidum) occurring in Australia and Italian rye grass (Lolium multiflorum) in Oregon (Stanger and Appleby, 1989).
| Table 3: | Parameter estimates of the susceptible and resistant populations survival as a percentage of untreated control, 4 weeks after spraying fenoxaprop-p-ethyl. Data were fitted according to the non-linear regression model: Y = k/(1+ebgxb)+d |
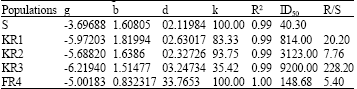 | |
| Parameters definition as in Table 2 | |
Heap et al. (1993) reported that A. fatua population UM1 was 150 times more resistant sethoxydim than susceptible populations. Figure 2 show the effect of difference fenoxaprop-p-ethyl concentrations on the survival of resistant and susceptible populations. In the resistant populations, four levels of response to fenoxaprop-p-ethyl were evident:
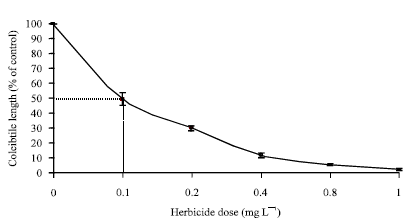 | |
| Fig. 3: | Effect of different fenoxaprop-p-ethyl concentrations on coleoptile’s length of susceptible population (S), as a percentage of untreated controls |
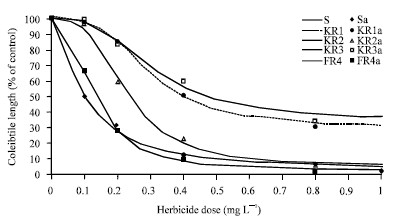 | |
| Fig. 4: | Effect of different fenoxaprop-p-ethyl concentrations on coleoptile’s length of susceptible (S) and resistant (KR1, KR2, KR3) populations as a percentage of untreated controls, 7 day after herbicide application. Symbols and lines represent actual and estimated response, respectively |
KR3> KR1> KR2> FR4 (Table 2). These results were confirmed with relationship between survived plants in these populations and fenoxaprop-p-ethyl doses (Fig. 2 and Table 3).
Seed bioassay
Discriminating dose experiment: At 7 DAHA, fenoxaprop-p-ethyl at 0.1 mg L-1 -1 inhibited population S Coleoptile’s length by 50% (Fig. 3). Thus, 0.1 mg L-1 was chosen as the discriminating dose.
Results of statistical analysis 7 DAHA showed that fenoxaprop-p-ethyl significantly affected the populations coleoptile’s length (Table 1). Results showed that KR1, KR2, KR3 and FR4 germinated almost completely which is consistent with our finding in whole plant assay. Thus, these populations exhibited resistance to fenoxaprop-p-ethyl but the other populations were susceptible, although susceptibility of some populations was lower than S population. Bena Kashani et al. (2006) also observed resistance to clodinafop-propargyl in Khuzestan wild oat populations.
Dose-response experiments: Result of next experiment indicated that KR1, KR2, KR3, FR4 were resistant to fenoxaprop-p-ethyl. The response of resistant and susceptible (S) populations to increasing dose of fenoxaprop-p-ethyl is shown in Fig. 4. Effect of fenoxaprop-p-ethyl doses on coleoptile’s length was visible as soon as germination was initiated and after 7 days there were large difference between the KR1, KR2, KR3, FR4 and S biotypes. Detailed dose response curves have confirmed these observations (Table 4). The effective concentration of herbicide causing 50% reduction in coleoptile’s length (ID50) was estimated from the dose-response curves (Tale 6). In these experiments similar to whole plant assay, the ranking of populations resistance ratio was KR3> KR1> KR2> FR4. FR4 is considered as a low level resistant population because its R/S ratio was not much greater than 1. Population resistance levels that were obtained at seed bioassay were lower than those obtained at whole plant assay. Tal et al. (2000) stated that although the seed bioassay seems to be less accurate compared to the whole plant assay (lower R/S values) it is.
| Table 4: | Parameter estimates of the coleoptile’s length of susceptible and resistant populations coleoptile’s length as a percentage of untreated controls, 7 day after fenoxaprop-p-ethyl application. Data were fitted according to the non-linear regression model: Y = k/(1+ ebg xb)+d |
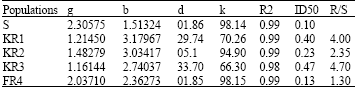 | |
| Parameters definition as in Table 2 | |
A reliable method for identifying populations of grass species resistant to ACCase inhibiting herbicides. Researcher confirmed the utility of the seed bioassay procedure for identifying ACCase inhibitor resistant wild oat populations, by testing appropriate concentrations of fenoxaprop-p and sethoxydim (Murray et al., 1996). The seed bioassay technique is a simple, comparatively quick and inexpensive, reliable and is particularly useful for routine screening of a large number of susceptible or resistant populations (Heap, 1994). The close association between the results from two tested methods may be represented a similar response to the same physiological-biochemical trait-resistance to ACCase inhibitors (Tal et al., 2000).
CONCLUSIONS
Results show that KR1, KR2, KR3, populations collected from Khuzestan province and FR4 population collected from Fars province have been confirmed to be resistant to fenoxaprop-p-ethyl. The results and conclusions using seed bioassay were generally similar to those based on whole plant assay. The rapid and accurate identification of resistant weed populations through use of seed bioassay system will assist in determining the nature and extent of the problem of ACCase inhibitor resistance on the Iran wheat fields. Alternative and effective weed management practices can then be implemented before the problem becomes unmanageable.
REFERENCES
- Beckie, H.J., L.F. Friesen, K.M. Nawolsky and I.N. Morrison, 1990. A rapid bioassay to detect trifluralin-resistant green foxtail (Setaria viridis). Weed Technol., 4: 505-508.
Direct Link - Beckie, H.J., I.M. Heap, R.J. Smeda and L.M. Hall, 2000. Screening for herbicide resistance in weeds. Weed Technol., 14: 428-445.
CrossRefDirect Link - Brain, P. and R. Cousens, 1989. An equation to describe dose responses where there is stimulation of growth at low doses. Weed Res., 29: 93-96.
CrossRefDirect Link - Do-Soon, J., C. Caseley, P. Brain, C.R. Riches and B.E. Valverde, 2000. Rapid detection of propanil and fenoxaprop resistance in Echinochloa colona. Weed Sci., 48: 695-700.
Direct Link - Heap, I.M., B.G. Murray, H.A. Loeppky and I.N. Morrison, 1993. Resistance to aryloxyphenoxypropionate and cyclohexanedione herbicides in wild oat (A. fatua). Weed Sci., 41: 232-238.
Direct Link - Murray, B.G., L.F. Friesen, K.J. Beaulieu and I.N. Morrison, 1996. A seed bioassay to identify acetyl-COA carboxylase inhibitor resistant wild oat (Avena fatua) populations. Weed Technol., 10: 85-89.
Direct Link - Sandral, G.A., B.S. Dear, J.E. Pratley and B.R. Cullis, 1997. Herbicide dose rate response curve in subterranean clover determined by a bioassay. Aust. J. Exp. Agric., 37: 67-74.
Direct Link - Tal, A., E. Kotoula and B. Rubin, 2000. Seed bioassay to detect grass weeds resistance to acetyl coenzyme-A inhibiting herbicides. Crop Prot., 19: 467-472.
Direct Link