
A. A. Mohammadpour
Department of Basic Sciences, Faculty of Veterinary Medicine, Ferdowsi University of Mashhad, P.O. Box 1793, Mashhad, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 4 | Page No.: 673-675







ABSTRACT
In this project, for measuring and comparing dimensions of ovary and ovarian follicles between Lori-Bakthiari sheep and native goat, eighty ovaries of sheep and goat, aged 1-2 years, from Shahrekord abattoir were collected. After weighing the ovaries, some factors such as: length, width and thickness of left and right ovaries by caliper device were measured. For histological studies, after tissue preparation and staining with H and E, diameter of follicles, thickness of follicular layers (granulosa, theca interna and theca externa) by micrometry methods were measured. By analyzing data with t-test and comparing them between sheep and goat we concluded that, there were significant difference between weight and dimensions of ovaries in sheep and goat. These parameters in goats were larger than sheep. Microscopical studies revealed that, the mean thickness of theca externa of graafian follicle in left ovary of sheep was more than right. The mean diameter of oocyte in secondary follicle of goat in left ovary was more than right. Also in goat, the number of layers of cumulus oophorus around corona radiata was more than sheep.
PDF Abstract XML References Citation
How to cite this article
A. A. Mohammadpour, 2007. Comparative Histomorphological Study of Ovary and Ovarian Follicles in Iranian Lori-Bakhtiari Sheep and Native Goat. Pakistan Journal of Biological Sciences, 10: 673-675.
DOI: 10.3923/pjbs.2007.673.675
URL: https://scialert.net/abstract/?doi=pjbs.2007.673.675
DOI: 10.3923/pjbs.2007.673.675
URL: https://scialert.net/abstract/?doi=pjbs.2007.673.675
INTRODUCTION
Quantitative aspects of follicle growth have been studied in sheep (Draincourt et al., 1993; Draincourt, 1991), bovine (Singh and Adames, 2000), mouse (Satosi and Motoaki, 2004) and wapiti (McCorkel et al., 2004). In all mammals, follicles begin to grow from a pool of primordial follicles constituted early in life, continuously throughout the life of the female. Primordial follicles are the least developed and most numerous follicles of the ovary. Each consists of a primary oocyst surrounded by a layer of simple squamous follicular cells. In late stages it developed to primary, secondary and graffian folliclee, respectively (Banks, 1993). Sheep and goat play a major role in modern agriculture and their size and physiology provide an appropriate model to study a variety of mammalian biological functions, including reproduction, embryology and fetal development. Sheep have several heritable traits of economic interest, such as those affecting fertility. These genetic variations occur in different breeds and provide interesting models to study the mechanisms regulating the formation and development of ovarian follicles.
Knowledge of ovarian follicular growth and maturation in sheep and goat is still limited. In spite of their great economic importance, detailed comparative studies have not been made of morphological and histological aspects of follicular growth in the sheep and goat. The comparison of follicle growth in goat and sheep will help in understanding the genetic differences. The results will be compared and contrasted not only among sheep and goat but also other mammalian species.
MATERIALS AND METHODS
Eighty ovaries of sheep and goat, aged 1-2 years, in summer season from Shahrekord abattoir were collected. They were immediately brought to laboratory in ice, washed throughly with 0.9% saline. In each specimen, some parameters such as: weight, length, width and thickness of left and right ovaries were measured by calliper device. For histological studies, after tissue preparation and staining with H and E, they were recognized and classified into the following categories:
A-Primordial follicle
B-Primary follicle (single layer, multi layer)
C-Secondary follicle (C-shaped antrum)
D-Graafian follicle
Then, diameter of follicles, thickness of follicular layers (granulosa, theca interna and theca externa) by micrometry methods were measured. In results, all of parameters with student t-test analysed and between sheep and goat were compared.
RESULTS
Results obtained from dimensions of ovaries in sheep and goat revealed that, all of parameters in goat was more than sheep and different was statistically significant (p<0.05) (Table 1).
| Table 1: | Comparative dimensions of ovary between sheep and goat |
 | |
| * Significant with p<0.05, values are shown in mean±SD | |
| Table 2: | Comparative dimensions of left and right ovary in sheep and goat |
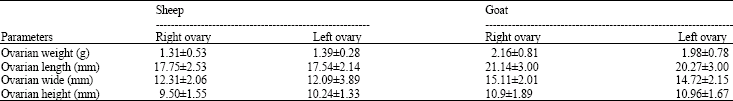 | |
| Values are shown in mean±SD | |
| Table 3: | Comparing Statistical analysis of ovarian follicles between sheep and goat |
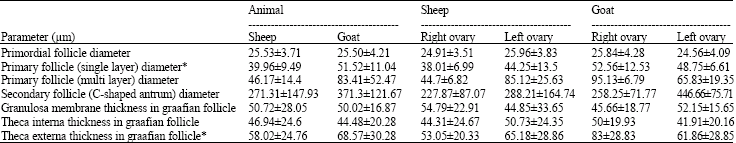 | |
| * Significant with p<0.05, values are shown in mean±SD | |
By comparing the dimensions of ovaries between left and right ovaries in sheep and goat, morphological parameters in both animals were same and there was no significant difference (Table 2). In histological studies, follicular layers of ovaries in sheep and goat by micrometry method were measured. After comparing measurements between left and right ovaries in two animals we concluded that, there is a significant difference (p<0.05) in theca externa thickness of graafian follicle In sheep and in left ovary (65.18±28.86 μm) was more than right (53.05±20.33 μm). Also in primary follicle (single layer) diameter of animals the difference was detectable and in goat (51.52±11.04 μm) larger than sheep (39.96±9.49 μm) were observed and the difference was statistically significant. There was no significant difference between other measurements (Table 3).
DISCUSSION
In this study, the differences between the mean follicle diameters except primary follicle (single layer) in goat and sheep was statistically non significant (p<0.05). Some of morphological factors of ovary in goat was larger than sheep. This difference might also be due to some qualitative and quantitative differences in circulating hormones of the two animals. There may be some differences in ovarian receptors for hormones.
In response to periodic hormonal stimulation, growth is initiated in some of the primordial follicles. The major developmental events that occur in the primary follicle include FSH receptor expression and oocyte growth and differentiation. Further growth results in the formation of a secondary follicle with a C-shaped antrum. Its follicular cells are now called the membrana granulosa. A sheath of stromal cells, the theca folliculi, forms around the follicle. The theca differentiates into a cellular, vascular inner layer, the theca interna and an outer, connective tissue layer, the theca externa. Continued growth results in the formation of a large tertiary (Graafian) follicle whose oocyte is surrounded by a multi layer of membrana granulosa cells, the cumulus oophorus. The columnar cells of the innermost portion of the latter constitute the corona radiata, which is separated from the oocyte by the zona pellucida (Banks, 1993; Monniaux et al., 1997). In most domestic animals the later stages of follicles development occurs in a wave-like pattern during oestrous cycles (cattle, sheep, goats, horses and buffalo) or periods of reproductive activity (llamas and camels). A follicle wave is the organized development of a cohort of gonadotrophin-dependent follicles all of which initially increase in size, but most of which subsequently regress and die by atresia (subordinate follicles). The number of remaining (dominant) follicles is specific to the species and is indicative of litter size. Follicle waves developed during both luteal and follicular phases and it is the dominant follicle(s) of the last follicular wave that ovulates. However, there are cases where dominant follicles from the last two follicle waves can ovulate (sheep and goats). There are exceptions to the organized wave-like pattern of follicle growth where follicle development is apparently continuous (pigs and chickens). In these animals many follicles develop to intermediate diameters and at specific times follicles that are destined to ovulate are selected from this pool and continue growing to ovulation. Understanding the pattern of follicle development in different species is increasingly important for designing improved methods to manipulate reproduction in domestic animals (Evans, 2003).
In another study was designed to quantitatively characterize the histomorphological attributes of dominant and subordinate follicles in relation to follicular wave dynamics. Heifers were examined daily using ultrasonography to record the growth of individual follicles from 2 days before ovulation until the day of ovariectomy. In results, follicular status, described by ultrasonography, was associated with quantitatively distinct histomorphological characteristics. Morphometric changes in the dominant follicle during immature, mature and post-mature phases were similar to, but occurred later than, those of subordinate follicle (Singh and Adames, 2000).
Ovarian follicles of sheep were studied by electron microscopy from primordial up to a stage shortly before ovulation. The follicular cells of primordial follicles were undifferentiated and rested on a thick basal lamina. In secondary follicles the endoplasmic reticulum had proliferated, forming an anastomosing network. In early antral and antral follicles the endoplasmic reticulum was composed of short cisternae, the mitochondria had enlarged and gap junctions were first observed. In late antral follicles gap junctions were frequent. In the cumulus the glycogen was associated with electron lucent areas, whereas in the granulosa it was invariably associated with membranes (Cran et al., 1979).
An improved knowledge of these factors will provide helpful clues in which local factors need to be manipulated to alter ovarian function and here, production of quality oocytes after superovulation for embryo transfer, polyembryony and genetic engineering technologies in small ruminants. Such technologies are better developed for cattle (Lauria and Gandolfi, 1992).
REFERENCES
- Draincourt, M.A., 1991. Follicular dynamics in sheep and cattle. Theriogenology, 35: 55-79.
Direct Link - Evans, A., 2003. Characteristics of ovarian follicle development in domestic animals. Repro. Domestic Anim., 38: 240-246.
CrossRefDirect Link - McCorkel, R.B., L. MacDougall and G.P. Adams, 2004. Ovarian follicle development in wapiti (Cervus elaphus) during the anovulatory season. Theriogenology, 61: 473-483.
Direct Link - Monniaux, D., C. Huet, N. Besnard, F. Clement and M. Bosc et al., 1997. Follicular growth and ovarian dynamics in mammals. J. Reprod. Fertil. Suppl., 51: 3-23.
Direct Link - Satosi, K. and U. Motoaki, 2004. Histological analysis of the critical point in follicular development in mice. Reprod. Med. Biol., 3: 141-145.
Direct Link - Singh, J. and G.P. Adams, 2000. Histomorphometry of dominant and subordinate bovine ovarian follicles. Anat. Rec., 258: 58-70.
Direct Link