
N. Sinsiri
Department of Agricultural Technology, Mahasarakham University, Mahasarakham 44000, Thailand
S. Laohasiriwong
Department of Agronomy, Khon Kaen University, Khon Kaen 40002, Thailand
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 1 | Page No.: 65-71







ABSTRACT
This experiment was carried out in the dry season (February-April, 2004) at the Experimental Farm, Mahasarakham University, Northeast Thailand to investigate the effect of different rates of indole-3-acitic acid (IAA) growth regulator in inducing root formation of detached leaves of MSU1, KVC#7 and IT84E-1-108 cowpea cultivars under tunnel conditions. The experiment was laid in a strip plot design with four replications and each replication was duplicated four times for four sampling periods. IAA levels used were 0, 250, 500, 750, 1,000, 1,500 and 2,000 mg L-1 of distilled water, thus the experiment consisted of 21 treatments. The results showed that root length, number of both roots and root hairs were highly affected by IAA treatments and the best IAA level was found with level 3 (500 mg L-1). The effects due to IAA levels and cultivars were highly significant and the effects due to an interaction between factors A (cultivars) and B (IAA levels), in most cases, were highly significant. Detached leaf technique with the use of IAA growth regulator could be considered as one of the most appropriate techniques for the preparation of plant samples for the test on disease resistance of cowpea cultivars.
PDF Abstract XML References Citation
How to cite this article
N. Sinsiri and S. Laohasiriwong, 2007. Effects of Different Rates of Indole-3-Acitic Acid on Root Formation of Detached Leaves of Cowpea (Vigna unguiculata L. Walp). Pakistan Journal of Biological Sciences, 10: 65-71.
DOI: 10.3923/pjbs.2007.65.71
URL: https://scialert.net/abstract/?doi=pjbs.2007.65.71
DOI: 10.3923/pjbs.2007.65.71
URL: https://scialert.net/abstract/?doi=pjbs.2007.65.71
INTRODUCTION
Within the past few decades, the cultivation of a number of leguminous crops in Thailand has been facing with many problems in obtaining high crop yields due to the spread out of the various types of diseases cause by microorganisms such as bacteria, fungi and others where they obviously damaged crop yields (Frey et al., 1977; Allen et al., 1998; Awurum, 2000; Emechebe and Lagoke, 2003; Sinsiri et al., 2006). A rapid assessment on the disease intensity when the crops are infected could be one way to cope up the problems and at the same time workers could be able to select, which crop cultivars are to be chosen for further crop cultivation and breeding programme. Growing crops for disease testing under field conditions may require many more months to complete the work and the expenses could be relatively high such as with the case of soybean (Glycine max) it may take 2-3 months to complete the work. Thus experiments on detached leaf technique may be of a significant value for a rapid assessment on disease infection of the crops since the plant samples are allowed to produce roots under water solutions within a short period of time. The use of detached leaf technique for the induction of roots has been used by a number of workers, e.g., Liu et al. (1996) showed that hypocotyls of naphthaleneacetic acid (NAA) treated cuttings of soybean (Glycine max) grew significantly higher numbers of adventitious roots with an increase in endogenous IAA levels where it corresponded to a decrease in IAA oxidase activity. The increase in IAA levels with a decrease in IAA oxidase activity in NAA-treated tissues aided the induction of adventitious roots. They stated that the higher IAA levels accumulated in the plant tissues could have encouraged higher amounts of root formation. With another work on soybean experiment, Liu et al. (1998) reported that accumulation of endogenous indole-3-acetic acid (IAA) in soybean hypocotyls explants was attained during a period when adventitious root had formed. Their treatments include naphthaleneacetic acid (NAA) and indole-3-butyric acid (IBA). They suggested that the application of auxin (IBA and NAA) could have acted on polyamine synthase and IAA oxidase activity. Whilst Moncousin et al. (1989) with grape wine showed that node cuttings of an in vitro culture rooted in absence of any growth regulator. Before the onset of axillary bud, there were two peaks of ethylene production at 2 and 10-12 h well marked in the top and bottom portions of the cuttings for the former. Indole-3- acitic acid (IAA) increased in the basal portions of the cuttings from 4th h and cumulated at 24th h where ethylene induction of the first rise could have been initiated the sequences of reactions leading to root formation. Montoya et al. (1997) carried out experiments on common bean (Phaseolus vulgaris L.) in Puerto Rico and the Dominic Republic aiming to attain high resistant populations on the infection of web blight disease caused by Rhizoctonia solani Kuhn. With their screening method, they attained five lines of common bean crop, which possessed moderate to low levels of web blight infection in field experiments. Other workers have also applied a detached leaf screening technique for use in their crop selection programmes such as Bautista and Echavez-Badel (2000) on bean lines with respect to web blight disease, Takegami et al. (2004) also with bean plants on web blight. With another published data on peanut experiment, Favero et al. (2005) used 69 peanut species (Arachis sp.) for their experiment and they stated that detached leaf technique was an applicable technique in inducing root formation of detached leaves of several species of both hybrid and wild type of peanut. They stated that wild peanut species produce small amounts of seeds with few root meristems where the situations create some difficulties in evaluating cytogenetic characterization. The use of IAA in encouraging root formation for Aristolochia manshuriensis cuttings was also carried out by Hedman (2005). He stated that the best root formation of 70%was attained from cuttings treated with IAA at 500 mg L-1 after 34 weeks of a rooting period. Thus root development of the cuttings was encouraged by Went’s auxin, which was known as IAA, a plant growth regulator (Salisbury and Ross, 1992). For this work it was anticipated that the use of different rates of IAA for root induction under detached leaf technique could possibly provide adequate amounts of detached leaf samples for further experiments when the detached leaves were able to produce some large amounts of roots and able to attain a normal survival. Thus an inoculation of some specific pathogenic diseases to the detached leaf samples could be carried out. In carrying out the experiment as such it could possibly assist scientists, particularly plant breeders to attain the best dominant genes of the crops where the plant species possess high resistant property against diseases and thus the outstanding plant species could be selected for further breeding programme.
MATERIALS AND METHODS
This experiment was conducted in a closed system glasshouse at the Department of Agricultural Technology, Mahasarakham University, Mahasarakham Province, Thailand during February-April 2004 to investigate the effect of different rates of indole-3-acetic acid (IAA) on rooting induction of detached `leaves of cowpea (Vigna unguiculata L. Walp). The experiment was laid in a strip plot design with four replications. MSU1, KVC#7 and IT84-1-108 cowpea cultivars were used as vertical strip (Factor A). IAA rates used were 0 (level 1), 250 (level 2), 500 (level 3), 750 (level 4), 1,000 (level 5), 1,500 (level 6) and 2,000 mg L-1 (level 7) of distilled water and they were used as horizontal strip (Factor B), thus the experiment consisted of 21 treatments. Each treatment had 4 replications and each replication was duplicated 4 times for sub-samples where they were used for sampling periods at 7 days interval starting from day 7 up to day 28, i.e., they were altogether 4 sampling periods, hence the experiment consisted of 336 units of detached leaves. A preparation for plant samples for use in the experiment was carried out with the use of 220 plastic trays, each tray has a diameter of 26 cm and each of them contained an amount of compost of 6 kg for use in germinating and growing of the cowpea plants. Four to five seeds of each cowpea cultivars were sown into the trays and then watered with an equal amount of tapped water to each pot to reach nearly a field capacity. All of the trays were placed under the glasshouse to avoid disease contamination. Three days after germination, seedlings were thinned out leaving only two seedlings/pot. Daily watering of the pots was carried out early in the morning. At 28 days after germination, the forth leaf of each plant (being counted from top) was chosen with a plastic marker on it and then the two leaves adjacent to the chosen leaf were taken away. This was carried out in order to maintain high water turgidity in cells of the chosen leaves. After the unwanted two leaves were removed for 2 days then the chosen leaves were cut out from each cowpea plant, i.e., 30 days after emergence. Three hundred and thirty six transparent plastic containers (PVC containers) with diameter of 7.5 cm and 11.5 cm long were used. Each plastic container has a rounded hole at the base of the container with a diameter of 3 mm (top end opened) and then each plastic container was filled up with 250 g of sand particles. Before filling up the sand particles into the containers, the sand particles were sterilized under a pressure of 15 lbs/sq. inch at 121°C for 30 min and then allowed to cool down in the glasshouse. Twenty eight trays filled up with distilled water at a height of 1 cm were used. Each tray had 12 plastic containers submerged under their respective IAA treatments. This aimed to attain some considerable amounts of capillary distilled water to be absorbed through sand particles in the pipes throughout the rooting period. The amounts of distilled water were maintained at 1 cm high (measured from the inner basement of the trays) throughout the experimental period. Petiole of each chosen detached leaf was cut out in distilled water to avoid air bubbles in the wounded petioles. Each petiole was cut out again at 5 cm long (measured from the edge of each leaf) and then each of them (petiole) was immediately submerged into their respective treatments of IAA solutions for 2 min and then buried into sand particles of each plastic container to the depth of approximately 3-4 cm. All of the trays of the plant samples were placed into a closed system plastic tunnel with an area of 12 m2 (3x4 m). The roofing height of the tunnel was 2.5 m above ground level. The tunnel was covered with transparent plastic films where approximately 50% of natural light intensity from the sun could be able to pass through. The relative humidity level of approximately 100% inside the tunnel was maintained throughout the experimental period with the use of mist spraying of distilled water three times daily to avoid the lost of water in leaves through evapo-transpiration. Daytime temperature in the tunnel was maintained at approximately 33°C with the use of outlet plastic films and nighttime temperature was not below 28°C. The plastic pipes in the trays with plant samples were placed under controlled temperature and relatively humidity for 1 week then the plastic sheets surrounded the tunnel were removed when roots had developed at each cutting of the leaf petioles.
The following sampling parameters on root development were used for each detached leaf sample, i.e., root length, number of roots and number of root hairs. This was carried out at 7 day intervals soon after the plant samples were placed under the tunnel starting from day 7 up to day 28, i.e., 4 sampling periods were carried out. The results on number of roots on days 7 and 14 are included in this study, whilst the data taken at days 21 and 28 are excluded because they gave a similar trend due to treatments as that of days 7 and 14. The data on number of root hairs taken at days 14 and 21 are excluded since, in most cases, both provided a similar trend to day 28. The attained results were statistically analysed using MSTAT-C statistical methods (Nissen, 1989).
RESULTS
Root length: With MSU1 cowpea cultivar, the results derived from two-way analysis on root development of detached leaves at day 7 revealed that an increase in IAA levels significantly increased root length up to level 4 (750 mg L-1) although further increases in IAA levels gave a similar significant effect as that of level 4 (750 mg L-1) with mean values ranged from 9.85 to 72.50 cm for levels 1 and 7, respectively (Table 1). For KVC #7 cultivar, an increase in IAA levels significantly increased root length up to the highest level (level 7, i.e., 2,000 mg L-1) although level 3 was similar to level 4 with values ranged from 12.50 to 32.87 cm for levels 1 and 7, respectively. With IT84E-1-108 cultivar, an increase in IAA levels significantly increased root length up to level 3 (500 mg L-1) with values ranged from 7.65 to 58.25 cm for levels 1 and 3, respectively. The effect due to factor B (IAA levels) was highly significant where level 5 possessed the highest but similar to level 3 with values of 47.50 and 39.11 cm for levels 5 and 3, respectively. The effect due to factor A (cultivars) was also highly significant where MSU1 ranked the highest followed by IT84E-1-108 and KVC#7 with values of 52.50, 34.53 and 23.36 cm, respectively. For an interaction between the two factors, i.e., A (cultivars) and B (IAA levels), it revealed that an interaction between the two factors on root length was highly significant where MSU1 attained the highest followed by IT84E-1-108 and KVC#7 but KVC#7 was not statistically different from IT84E-1-108 cultivar.
At 14 days under the tunnel conditions, the results showed that an increase in IAA levels highly increased root length of all tested cultivars. The response of MSU1 cultivar to an increase in IAA levels was only up to level 3 (500 mg L-1) but this level was similar to level 6 and level 6 was similar to level 7 with mean values ranged from 116.00 to 241.50 cm for levels 1 and 3, respectively (Table 2). For KVC#7, an increase in IAA levels highly increased root length up to level 3 (500 mg L-1). Further increases in IAA levels, in most cases, were lower than level 3 although level 4 was similar to level 3 with mean values ranged from 100.25 to 232.25 cm for levels 1 and 3, respectively.
| Table 1: | A two-way analysis of root lengths (cm) in three cowpea cultivars as affected by IAA solution levels at day 7 under tunnel conditions |
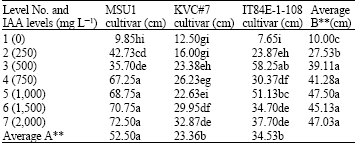 | |
LSD for factor A = 12.12 and for factor B = 9.09, AxB = 13.97, F-test for factor A = **, B = ** AxB = **, CV = 19.74%, Letter(s) within columns indicate Least Significant Differences (LSD) at p = 0.01 | |
| Table 2: | A two-way analysis of root lengths (cm) in three Cowpea cultivars as affected by IAA solution levels at day 14 under tunnel conditions |
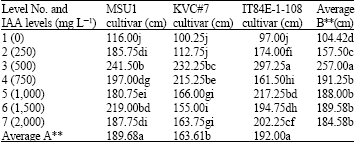 | |
LSD for factor A = 25.42 and for factor B = 20.19, AxB = 30.73, F-test for factor A = **, B = ** AxB = **, CV = 8.79%, Letter(s) within columns indicate Least Significant Differences (LSD) at p = 0.01 | |
With IT84E-1-108 cultivar, an increase in IAA levels highly increased root length of the detached leaves up to level 3. Further increases in IAA levels gave no consistent trend on root length due to treatments with mean values ranged from 97.00 to 297.25 cm for levels 1 and 3, respectively. The effect due to factor B (IAA levels) was highly significant where level 3 was the highest, whilst other higher IAA levels were not different from one another. With factor A (cultivars), the results revealed that MSU1 and IT84E-1-108 were similar and both ranked the highest and the lowest was found with KVC#7 with mean values of 189.68, 192.00 and 163.61 cm, respectively. For an interaction between the two factors, i.e., A and B, the interaction was highly significant.
At day 21, the results showed that an increase in IAA levels significantly increased root length of MSU1 cultivar where the increase was up to level 3 (500 mg L-1). Further increases in IAA levels gave a similar root length to that of level 3 with mean values ranged from 145 to 225.00 cm for levels 1 and 5, respectively (Table 3). For KVC#7, the results revealed that an increase in IAA levels significantly increased root length up to level 2 (250 mg L-1). Further increases in IAA levels gave a similar root length as that of level 2 except level 5 where the attained value was lower than level 4 with mean values ranged from 147 to 229.50 cm for levels 1 and 3, respectively. With IT84E-1-108 cultivar, the results showed that an increase in IAA levels significantly increased root length up to level 3 (500 mg L-1). Further increases in IAA levels gave a similar significant effect as that of level 3 with mean values ranged from 141.50 to 213.50 cm for levels 1 and 4, respectively. The effect due to factor B (IAA levels) was highly significant where level 4 was the highest but similar to other higher levels except level 5 where the root length was similar to level 2 but similar to most other levels except level 1. The effect due to factor A (cultivars) was significantly found where both MSU1 and KVC#7 were similar and ranked the highest, whilst IT84E-1-108 ranked the lowest. For an interaction between the two factors, i.e., A and B, it revealed that there was a highly significant interaction between the two factors.
With the results on day 28, it showed that an increase in IAA levels significantly increased root length of the detached leaves of the cowpea cultivars. For MSU1 cultivar, an increase in IAA levels significantly increased root length up to level 7 but similar to level 3 with mean values ranged from 149.50 to 227.25 cm for levels 1 and 7, respectively (Table 4). The root length of KVC#7 cultivar was significantly increased with an increase in IAA levels. The increase was up to level 3 and then a decrease although higher levels gave a similar root length due to treatments with values ranged from 213.25 to 272.25 cm for levels 1 and 3, respectively.
| Table 3: | A two-way analysis of root lengths (cm) in three cowpea cultivars as affected by IAA solution levels at day 21 under tunnel conditions |
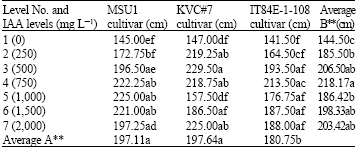 | |
LSD for factor A = 13.73 and for factor B = 22.55, AxB = 44.75, F-test for factor A = **, B = ** AxB = **, CV = 12.13 %, Letter(s) within columns indicate Least Significant Differences (LSD) at p** = 0.01 | |
| Table 4: | A two-way analysis of root lengths (cm) cowpea cultivars as affected by IAA solution levels at day 28 under tunnel conditions |
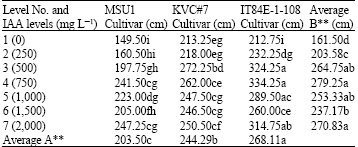 | |
LSD for factor A = 21.62 and for factor B = 29.28, AxB = 43.43, F-test for factor A = **, B = ** AxB = **, CV = 9.46 %, Letter(s) within columns indicate Least Significant Differences (LSD) at p = 0.01 | |
For IT84E-1-108 cultivar, the results showed that an increase in IAA levels highly increased root length up to level 3, further higher IAA levels gave a similar root length as that of level 3 except level 6 where root length was significantly lower than levels 4, 5 and 7. The effect due to factor B (IAA levels) was highly significant where level 4 was the highest but similar to level 3 with values of 279.25 and 264.75 cm, respectively. The effect due to factor A (cultivars) was also highly significant where IT84E-1-108 ranked the highest followed by KVC#7 and the least was found with MSU1 with values of 268.11, 244.29 and 203.50 cm, respectively. The effect due to factors A (cultivars) and B (IAA levels) interaction was highly significant.
Number of roots: With MSU1 cultivar at day 7, the results showed that an increase in IAA levels highly increased number of roots up to level 4 (750 mg L-1). Further levels gave a similar significant effect due to treatments as that of level 4 with mean values ranged from 8.25 to 41.00 for levels 1 and 4, respectively (Table 5). For KVC#7 cultivar, the results revealed that an increase in IAA levels highly increased number of roots up to level 7. There was no consistent trend due to an increase in IAA levels found with mean values ranged from 7.75 to 29.25 for levels 1 and 7, respectively.
| Table 5: | A two-way analysis of number of roots in three cowpea cultivars as affected by IAA solution levels, at day 7 under tunnel conditions |
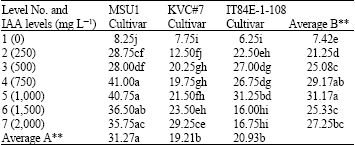 | |
LSD for factor A = 4.04 and for factor B = 3.59, AxB = 6.71, F-test for factor A = **, B = ** A x B = **, CV = 14.66 %, Letter(s) within columns indicate Least Significant Differences (LSD) at p = 0.01 | |
With IT84E-1-108 cultivar, the results showed that an increase in IAA levels highly increased number of roots up to level 5 and then a decline where all of the mean values became smaller than level 5 with mean values ranged from 6.25 to 31.25 for levels 1 and 5, respectively. With the effect due to factor B (IAA levels), the results showed that there was a highly significant effect due to IAA levels where level 5 was the highest, whilst the effect due to factor A (cultivars) was also highly significant where MSU1 ranked the highest followed by both KVC#7 and IT84E-1-108 cultivars where both gave a similar significant level. An interaction between factors A and B was also highly significant.
At day 14, the results showed that an increase in IAA levels highly increased number of roots of MSU1 cultivar up to level 6 but level 6 was similar to level 3 (500 mg L-1) with mean values ranged from 22.75 to 43.50 for levels 1 and 6, respectively (Table 6). With KVC#7, the results showed that an increase in IAA levels highly increased number of roots up to level 3. Further IAA levels gave significantly lower mean values than level 3 with mean values ranged from 19.75 to 44.00 for levels 1 and 3, respectively. For IT84E-1-108 cultivar, the results revealed that an increase in IAA levels highly increased number of roots up to level 3 and then a decline where all levels of IAA were similar and not statistically different from one another. The effect due to factor B (IAA levels) was highly significant where level 3 attained the highest with mean values ranged from 20.75 to 44.75 for levels 1 and 3, respectively. With factor A (cultivars), the results showed that there was a highly significant effect due to cultivars where MSU1 and IT84E-1-108 ranked the highest and the lowest was with KVC#7 with mean values of 37.27, 33.39 and 28.61, respectively. There was an interaction between factors A and B where they gave significant effect due to both cultivars and IAA levels.
Number of root hairs: The counting for number of root hairs was possible at days 14, 21 and 28 but only the results at day 28 are included due to the reasons described earlier in materials and methods.
| Table 6: | A two-way analysis of number of roots in three cowpea cultivars as affected by IAA solution levels at day 14 under tunnel conditions |
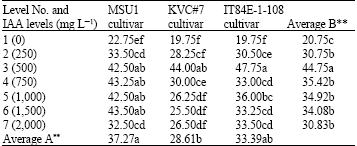 | |
LSD for factor A = 6.66 and for factor B = 7.23, AxB = 7.85, F-test for factor A = **, B = ** AxB = *, CV = 16.57 %, Letter(s) within columns indicate Least Significant Differences (LSD) at p** = 0.01,* = 0.05 | |
| Table 7: | A two-way analysis of number of root hairs in three cowpea cultivars as affected by IAA solution levels at day 28 under tunnel conditions |
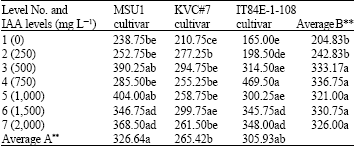 | |
LSD for factor A = 36.31 and for factor B = 65.23, AxB = 149.80, F-test for factor A = **, B = ** AxB = **, CV = 25.62 %, Letter(s) within columns indicate Least Significant Differences (LSD) at p ** = 0.01 | |
At day 28 the results showed that an increase in IAA levels highly increased number of root hairs of MSU1 cultivar but the increase was, in most cases, up to level 3 (500 mg L-1), further increases in IAA levels gave a similar significant level as that of level 3 with values ranged from 238.75 to 404.00 for levels 1 and 5, respectively (Table 7). With KVC#7 cultivar, an increase in IAA levels highly increased number of root hairs up to level 6. The highest level of IAA (level 7) gave significantly higher number of root hairs than level 1 but similar to other levels with values ranged from 210.75 to 299.75 for levels 1 and 6, respectively. For IT84E-1-108 cultivar, an increase in IAA levels highly increased number of roots up to level 4 where level 4 was similar to level 3. Further increases in IAA levels gave a similar number of roots as that of level 3 with values ranged from 165.00 to 469.50 for levels 1 and 4, respectively. The effect due to cultivars (factor A) on IAA levels was highly significant where MSU1 gave the highest but similar to IT84E-1-108 and IT84E-1-108 was similar to KVC#7. The effect due to IAA levels (factor B) was also highly significant where level 3 was the highest but similar to other higher IAA levels. An interaction between factors A and B was also highly significant.
DISCUSSION
It is well established that most of cereal crops being cultivated under field conditions are susceptible to the infection of some pathogenic diseases cause by different types of pathogenic organisms such as bacteria, fungi and viruses (Galvez et al., 1989; Godoy-Lutz et al., 1996; 1998; Takegami et al., 2004). The infection of such pathogenic diseases in crops has tremendously caused poor annual production of crop yields. Thus with this investigation the preparation for cowpea detached leaf samples with the use of IAA solutions where the plant samples were allowed to produce roots was successfully attained. The results showed that root lengths of cowpea detached leaves of MSU1, KVC#7 and IT84E-1-108 cultivars were highly induced by IAA levels throughout the experimental period commencing from the first interval after buried in sand particles (control treatment) with the use of distilled water added up to the trays. Nevertheless, at day 7 the responses of the three cultivars to IAA levels were not the same, i.e., the most appropriate level for MSU1 was with level 4 (750 mg L-1), level 6 for KVC#7 and level 3 for IT84E-1-108. Whilst at day 14, the most appropriate level of IAA for all cultivars was found with level 3 but at day 21, both KVC#7 and IT84E-1-108 responded well to level 2 whereas MSU1 responded well to level 3. However, at day 28 the effect due to IAA levels was relatively cleared, i.e., all of the cultivars responded most to level 3 (500 mg L-1), whereas the effect due to cultivars was not found. This could have been attributed to the longer duration (longer than 21 days after being placed under the tunnel) where the three cultivars had acquired some considerable amounts of roots. The differences found among the three cultivars at day 7 up to day 14 may be attributable to the differences in genetic traits where the leaves of each cultivar may have possessed different rates of photosynthetic activities but became similar when they reached the forth interval (28 days of rooting period). The results on root lengths indicated that it is possible to inoculate pathogenic organisms to the plant samples even at the beginning of the second week since some considerable amounts of roots had produced. Therefore, an assessment on the resistance to diseases of the cultivars could be examined most rapidly even during the second week of the experimental period. Thus detached leaf technique could be an appropriate technique for plant disease assessment. Leaf senescence is an important factor to be encountered hence the disease inoculation should be carried out soon after roots had developed, i.e., at least during the fifth day of rooting period.
With number of roots of the cowpea cultivars at day 7 after submerged in the sand particles, the results showed that all of the cultivars highly responded to IAA levels where, in most cases, level 5 was the highest. The differences among the cultivars may be attributable to genetic traits where efficacy in photosynthetic activity and amounts of carbohydrate reserved may not be the same. However, the results taken at day 14 were relatively cleared that is all cultivars responded well to level 3 (500 mg L-1). Thus this level of IAA could be considered as the most appropriate level for use in inducing some considerable amounts of root of the cowpea plants. The results on the amounts of root hairs at day 28 confirm this level of IAA to be used for further experiments. The results also indicated that detached leaf technique being used in inducing root formation is workable hence the plant samples could be ready for use for any inoculation of pathogenic organisms for further experiments. These attained results are comparable to the work on root formation of the cuttings carried out by Liu et al. (1996 and 1998) with soybean; Moncousin et al. (1989) with grape cuttings, Bautista and Echavez-Badel (2000) with bean lines; Takegami et al. (2004) also with bean plants and also Alessandra et al. (2005) with 69 peanut species. Therefore, detached leaf technique could be one of the most appropriate techniques for use in selecting for high resistant cowpea cultivars against pathogenic diseases when tested under tunnel conditions and this Went’s IAA growth regulator (Salisbury and Ross, 1992) can be successfully used in inducing root formation of cowpea detached leaf experiments but the most suitable level could be only up to level 3 (500 mg L-1).
ACKNOWLEDGMENTS
The authors wish to thank Mahasarakham University Financial Office for financial assistance. Thanks are also due to staff members of the Faculty of Technology, Mahasarakham University for their kind assistance when this work was carried out.
REFERENCES
- Favero, A.P., L.L. Nass, A. Cargnin and E.L. Neto, 2005. Rooting performance from leaf petioles of accession and hybrids of wild Arachis species. Sci. Agric. (Piracicaba, Braz.), 62: 62-68.
Direct Link - Godoy-Lutz, G., J. Arias, J.R. Steadman and K.M. Eskridge, 1996. Role of natural seed infection by the web blight pathogen in common bean seed damage, seedling emergence and early disease development. Plant Dis., 80: 887-890.
Direct Link - Liu, Z.H., I.C. Hsiao and Y.W. Pan, 1996. Effect of naphthaleneacetic acid on endogenous indole-3-acetic acid, peroxidase and auxin oxidase in hypocotyl cuttings of soybean during root formation. Bot. Bull. Acad. Sin., 37: 247-253.
Direct Link - Liu, Z.H., W.C. Wang and Y.S. Yen, 1998. Effect of hormone treatment on root formation and endogenous indole-3-acetic acid and polyamine levels of Glycine max cultivated in vitro. Bot. Bull. Acad. Sin., 39: 113-118.
Direct Link - Moncousin, C., J.M. Favre and T. Gaspar, 1989. Early changes in auxin and ethylene production in vine cuttings before adventitious rooting. Plant Cell Tissue Org. Cult., 19: 235-242.
CrossRefDirect Link - Montoya, C.A., J.S. Beaver, R. Rodriguez, P.N. Miklas and G. Godoy-Lutz, 1997. Heritability of resistance to web blight in five common bean population. Crop Sci., 37: 780-783.
Direct Link - MSTAT Development Team, 1989. MSTAT user's guide: A microcomputer program for the design, management and analysis of agronomic research experiments. Michigan State University East Lansing, USA pp: 1-152.
Direct Link - Sinsiri, N., S. Laohasiriwong, S. Jogloy, B. Toomsan and W. Saksirirat, 2006. A varietal screening of cowpea cultivars (Vigna unguiculata) for a high resistance to Pseudocercospora cruenta (Sacc.) Deighton in Northeast Thailand. Pak. J. Biol. Sci., 9: 641-648.
CrossRefDirect Link