
Masoud Sheidai
Department of Biology, Shahid Beheshti University, Tehran, Iran
Saeideh Attaei
Department of Biology, Shahid Beheshti University, Tehran, Iran
Masoomeh Khosravi-Reineh
Department of Biology, Shahid Beheshti University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 4 | Page No.: 622-626







ABSTRACT
Numerical taxonomic studies were performed on 57 populations of 15 Stipa species belonging to 5 different sections, concerning intra-population and inter-populations variations as well as inter-specific relationships. The most variable morphological characters useful in the species delimitation were also determined. The species studied also differed significantly in most of the quantitative characters studied and the mean of such characters may be of use in the species delimitation. The clustering showed distinctness of the species studied as the populations of each species are placed close to each other and separate from the other species.
PDF Abstract XML References Citation
How to cite this article
Masoud Sheidai, Saeideh Attaei and Masoomeh Khosravi-Reineh, 2007. Numerical Taxonomy of Stipa (Poaceae) Species in Iran. Pakistan Journal of Biological Sciences, 10: 622-626.
DOI: 10.3923/pjbs.2007.622.626
URL: https://scialert.net/abstract/?doi=pjbs.2007.622.626
DOI: 10.3923/pjbs.2007.622.626
URL: https://scialert.net/abstract/?doi=pjbs.2007.622.626
INTRODUCTION
The genus Stipa L. (Poaceae) is comprised of 300-400 species throughout the world, with about 90-100 species distributed in the old world (Freitag, 1985; Barkworth and Everett, 1987). Discrepancy about the umber of Stipa species is due to taxonomic problems existing in this genus (Vázquez and Devesa, 1997).
The Stipa species distributed in various regions of the country are among important forage plants of Iran. The number of Stipa species growing in Iran varies from 6 to 18 according to different authors (Parsa, 1950; Bor, 1970; Freitag, 1985). Although the available literature dealing with systematic and biosystematics of Stipa species indicate the importance of these taxa (Freitag, 1985; Tzvelev, 1989), no report is available on the biosystematics of Stipa species and populations from Iran and only recently some basic information about cytogenetics of some of the species has been provided (Sheidai and Attaei, 2005). Therefore the present study considers the numerical taxonomic study of 57 populations belonging to 15 Stipa species growing in Iran, trying to reveal the inter-population morphological variations and inter-specific interrelationships.
MATERIALS AND METHODS
Morphometric studies were performed on 57 populations of 15 Stipa species belonging to 5 sections (Freitag, 1985) namely: (1)- Sect. Lasiagrostis (Link) Hackel, including St. caragana and St. haussknechtii, (2)- Sect. Aristella (Trin.) Hackel including St. bromoides (L. Doerf., (3)- Sect. Stipella Tzvelev, including St. parviflora and St. capensis, (4)- Stipa L., species group 1 (Eriostipa), including St. lessingiana Trin. and Rupr. and St. pennata L., species group 2 (Brevigeniculatae/unigenicuatae), including St. caucasica Schmalh. subsp. caucasica, species group 3 (Leiostipa), including St. capillata and (5) Sect. Barbatae Junge, including St. arabica Trin. and Rupr. (3 populations), St. hohenackerana Trin. and Rupr., St. ehrenbergiana Trin. and Rupr., St. gaubae Bor, St. iranica Freitag and St. holosericea Trin. (Table 1).
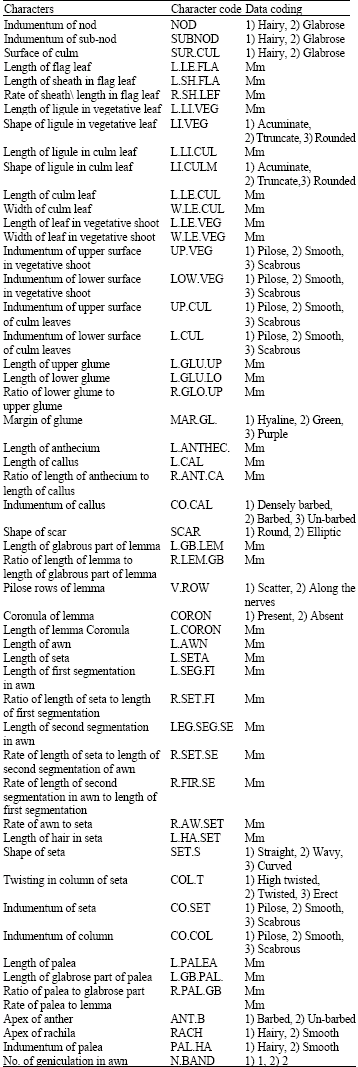
Three to five plants from each population were used for morphometric studies, from which 5-10 readings were done for characters studied. Details of the localities and the voucher numbers may be provided on request from the first author. Voucher specimens are deposited in TARI, IRAN and Herbarium of Shahid Beheshti University (HSB). In total 53 quantitative and qualitative morphological characters were studied (Table 1). Characters were selected based on those reported by Scagel and Maze (1984) and our own field observations.
In order to detect any significant difference in quantitative morphological characters among populations of each species and also among different species studied, analysis of variance (ANOVA) followed by the least significant difference (LSD) tests were performed.
In order to determine the species interrelationships, cluster analysis and principal component analysis (PCA) were performed (Ingrouille, 1986; Sheidai et al., 2002). For multivariate analyses the mean of quantitative characters were used, while qualitative characters were coded as binary/multistate characters. Standardized variables (mean = 0, variance = 1) were used for multivariate statistical analyses. The average taxonomic distance and squared Euclidean distance were used as dissimilarity coefficient in cluster analysis of morphological data (Podani, 2000). In order to determine the most variable morphological characters among the species studied, factor analysis based on principal components analysis (PCA) was performed (Podani, 2000). SPSS ver. 9 (1998) and STATISTICA ver. 5 (1995) were used for statistical analyses.
RESULTS AND DISCUSSION
The analyses showed that the range of size in quantitative characters studied is much different from what is given in different flora in the species descriptions Table 1. Therefore it is important to describe such characters after studying several populations of each species as performed in the present work. It is further supported by the results of ANOVA test as different populations of each species differed significantly in these quantitative characters.
The species studied also differed significantly in most of the quantitative characters as revealed by ANOVA and LSD tests. The mean of such quantitative characters may be useful for the species delimitation. This is partly supported by the clustering (Fig. 1) of the species studied based on merely quantitative morphological characters, as populations of some of the species are separated in distinct clusters including St. haussknechtii, St. iranica, St. hohenackerana, St. lessingiana and St. capensis (dented ha, I, ho, l and c respectively in Fig.1). However some populations of the species were mixed with the other species including St. arabica, St. parviflora and St. caragana in all of which only one population was separated and mixed with the other species.
The cluster analysis and PCA ordination of Stipa species studied based on both qualitative and quantitative characters produced similar results (Fig. 1-3). In the analyses based on all morphological characters, two major clusters or groups are formed. The first major cluster is comprised of two sub-clusters, in the first of which populations belonging to St. bromoides and St. caragana are placed close to each other. Bor (1970) in the Flora Iranica also considered these two species closely (morphologically) related to each other. Based on characters like presence of a rounded callus, straight awn measuring up to 35 mm and lemma with apical lobes, placed them in the sect. Lasiagrostis.However Freitag (1985) placed them in different sections.
| Table 1: | Morphological characters and their coding |
 | |
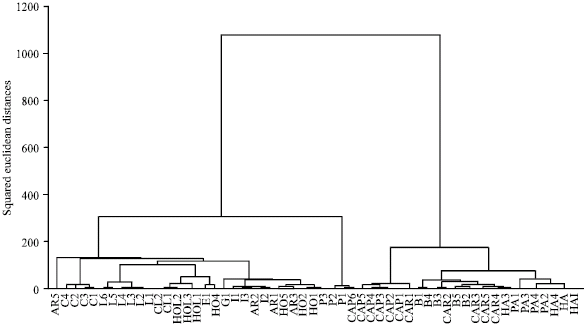 | |
| Fig. 1: | WARD clustering of Stipa species based on quantitative morphological characters. (Species code: CAR = St. caragana, HA = St. haussknechtii, B = St. bromoides, PA = St. parviflora, CAP = St. capensis, L = St. lessingiana, P = St. pennata; C = St. caucasica, CL = St. capillata, AR = St. arabica, HO = St. hohenackerana, G = St. gaubae, E = St. ehrenbergiana, I = St. iranica and HOL = St. holosericea) |
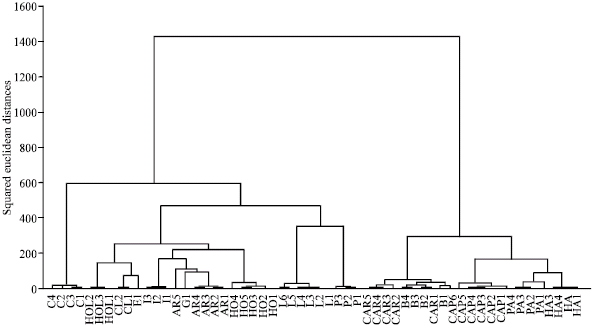 | |
| Fig. 2: | WARD clustering of Stipa species based on all morphological characters (species code as in Fig. 3) |
The second sub-cluster is comprised of three species, St. capensis, St. parviflora and St. haussknechtii in which, the last two species show more similarity and are placed close to each other. Bor (1970) in the Flora Iranica also considered the two species of St. haussknechtii and St. parviflora closely related to each other, while St. capensis is considered to be close to St. capillata.
Freitag (1985) based on the characters like palea being significantly (usually 2-4 x) shorter than lemma and the latter usually more or less constricted bellow the apex and by the extremely short upper lodicule, has placed St. capensis and St. parviflora in the sect. Stipella, which is supported by present analysis. But he considered St. haussknechtii close to St. caragana and placed them in the sect. Lasiagrostis, which is not supported by our analyses.
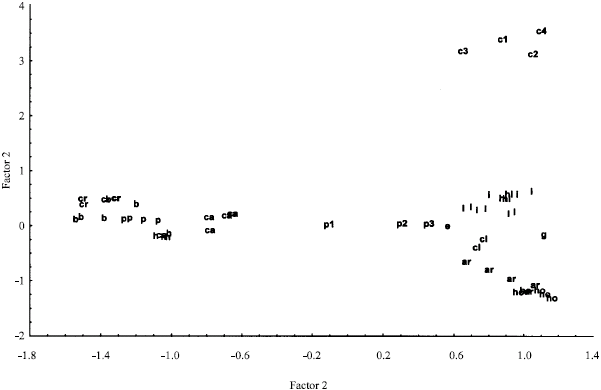 | |
| Fig. 3: | PCA ordination of the Stipa species based on all morphological characters (species code as in Fig. 1) |
The second major cluster is comprised of three sub-clusters. Populations of St. caucasica form the first sub-cluster/group much separated from the other species. Bor (1970) in the Flora Iranica considered this species close to St. gaubae, while Freitag (1985), considered this species close to St. capillata and St. lessingiana as the members of the section Stipa. However the present study dos not support these considerations.
The second sub-cluster is comprised of the species of St. holoserica, St. ehrenbergiana, St. iranica, St. arabica, St. gaubae and St. hohenackerana, supporting Freitag,s (1985) taxonomic treatment of the genus. Based on the presence of an ovary always with 3, more rarely 4, equal or unequal styles and stigmas he placed all these species in the sect. Barbatae.
The third sub-cluster is comprised of populations of St. lessingiana, St. pennata which, are along with St. capillata, are also grouped together in PCA plot, supporting Freitag,s (1985) taxonomic treatment of the genus. Based on the presence of a bigeniculate and hairy to plumose awn, he considered them close to each other in the species group 1 (Eriostipa) of the sec. Stipa.
However since Freitag (1985) stated that he is not sure about the position of St. gaubae, whether it is a member of sect. Stipa or sect. Barbata, cluster analysis was carried out only for the species present in these two sections only (Fig. 4). St. gaubae shows more similarity to St. arabica and St. hohenackerana of the sect. Barbata. Therefore the present numerical taxonomic study suggests inclusion of St. gaubae in the sect. Barbata.
In general, it seems that the present study partly supports Freitag,s (1985) taxonomic treatment of the genus Stipa. Therefore, it seems that more detailed studies are required to illustrate the species inter-relationships in the genus. In order to identify the most variable morphological characters among the species studied, PCA analysis was performed. The analysis revealed that the first 4 factors comprise about 67% of total variance. In the first factor with about 31% of total variance, characters like Apex of anther, length of upper glume, length of anthecium, length of awn, length of seta, ratio of seta to the first segment of awn, length of palea and ratio of palea to lemma possessed the highest positive correlation (>0.70). In the second factor with about 14% of total variance, characters like rachila, Length of ligule in culm leaf, indumentum of sub-nod and Indumentum of callus possessed the highest positive correlation (>0.50). Therefore these are the most variable morphological characters among the species studied. Qualitative and ratio characters identified in PCA along with those quantitative characters showing significant difference among the species studied (stated earlier) may be used in the Stipa taxonomy.
 | |
| Fig. 4: | WARD clustering of sections Stipa and Barbata species based on all morphological characters (species code as in Fig. 1) |
REFERENCES
- Sheidai, M., P. Koobaz, F. Termeh and B. Zehzad, 2002. Phenetic studies in Avena species and populations of Iran. I. R. J. Sci., 13: 19-28.
Direct Link - Sheidai, M. and S. Attaei, 2005. Meiotic studies of some Stipa (Poacea) species and populations in Iran. Cytologia, 70: 23-31.
Direct Link