
M. Sheidai
Department of Biology, Shahid Beheshti University, Tehran, Iran
M. Sottodeh
Department of Biology, Shahid Beheshti University, Tehran, Iran
B. Akbarei
Department of Biology, Shahid Beheshti University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 4 | Page No.: 553-560







ABSTRACT
Meiotic study was performed in 23 oil seed rape (Brassica napus) cultivars considering chiasma frequency and distribution as well as chromosomes pairing. All cultivars possessed n = 19 chromosome number (4x). Chromosome stickiness occurred in most of the cultivars from early prophase to late telophase-II leading to the formation of laggard chromosomes and micronuclei. The cultivars studied differed significantly in chiasma frequency and distribution as well as chromosomes association indicating their genomic differences. Cluster analysis and ordination based on principal components analysis grouped those cultivars showing meiotic similarities. Some of the cultivars showed the occurrence of B-chromosomes which significantly changed chiasma frequency and chromosome pairing in the cells possessing them. Unreduced meiocytes and pollen grains were observed in some of the cultivars due to cytomixis, anaphase failure and desynapsis.
PDF Abstract XML References Citation
How to cite this article
M. Sheidai, M. Sottodeh and B. Akbarei, 2007. Cytogenetic Variability in Several Oil Seed Rape Cultivars. Pakistan Journal of Biological Sciences, 10: 553-560.
DOI: 10.3923/pjbs.2007.553.560
URL: https://scialert.net/abstract/?doi=pjbs.2007.553.560
DOI: 10.3923/pjbs.2007.553.560
URL: https://scialert.net/abstract/?doi=pjbs.2007.553.560
INTRODUCTION
Oilseed rape/canola (Brassica napus L.) is among important oil producing plants cultivated extensively in Europe, China, North America and Iran. B. napus is an amphidiploid species with 19 pairs of chromosomes and has been evolved by crossing between B. campestris (2n = 2x = 20) and B. oleracea (2n = 2x = 18). Due to economic importance of oilseed rape, several B. napus cultivars have been introduced in Iran during the last decade and at present, germplasm evaluation as well as hybridization programs are in hand. Some basic cytogenetic information characteristics have already been reported in a few B. napus cultivars available in Iran (Sheidai et al., 2001a, b). The present work considers cytogenetic study of 23 unreported cultivars available in Iran considering the chromosome pairing and segregation, the occurrence and effects of B-chromosomes as well as unreduced gamete formation.
MATERIALS AND METHODS
Plants belonging to 23 B. napus L. cultivars were planted according to a randomized block design in the experimental field of Seed and Seedling Breeding Research Center, Karaj, Iran. For cytogenetical studies, flower buds were collected from 10 randomly selected plants from each cultivar. Fifty to hundred young flower buds were collected randomly during 9-12 A. M and fixed in glacial acetic acid: Ethanol (1:3) for 24 h which were then washed thoroughly and transferred to 85% ethanol until used (Sheidai et al., 2001a). Chromosome pairing and chiasma frequency was determined by using minimum 100 meiocytes showing diakinesis/ metaphase-I stages, while chromosome segregation was studied in minimum 500 anaphase-I and II stages. Pollen stainability as a measure of fertility was determined by staining minimum 1000 pollen grains with 2% acetocarmine: 50% glycerin (1:1) for about ½ h. Round/ complete pollens which were stained were taken as fertile, while incomplete/shrunken pollens with no stain were considered as infertile (Sheidai et al., 2003). Analysis of variance (ANOVA) followed by the least significant difference test (LSD) was performed on cytogenetic characteristics including chromosome pairing, chiasma frequency as well as distribution to indicate any significant difference among the cultivars studied (Sheidai et al., 2001a). Different methods of cluster analysis as well as ordination based on principal components analysis (PCA) was performed to identify the cultivars showing similarities in their meiotic characteristics (Sheidai et al., 2001a). For cluster and principal components analysis, standard values (mean = 0, variance = 1) were used. Squared Euclidean distance was use as a measure of similarity in cluster analysis (Sheidai et al., 2003). In order to detect the occurrence of unreduced (2n) pollen grains along with the normal (reduced = n) pollen grains, about 500 pollens were studied, some of which were sketched by the use of Camera Lucida and analysed by t-test. The statistical analyses used SPSS ver.9 (1998, SPSS Inc.) software.
RESULTS AND DISCUSSION
The B. napus cultivars studied and their meiotic characteristics are presented in Table 1 and 2 as well as Fig. 1-5. All cultivars possessed n = 19 (2n = 4x = 38) chromosome number. The highest value of total, terminal and intercalary chiasmata occurred in Chinese cultivar (36.48, 32.25 and 4.23, respectively), while the lowest values of total and terminal chiasmata occurred in SWO756 cultivar (21.39 and 20.68, respectively). The lowest value of intercalary chiasma occurred in the hybrid cultivar R.C-Chinese (4.21).
The highest value of ring bivalents occurred in Modena cultivar (14.94), while the lowest value occurred in SWO756 cultivar (3.09), which also possessed the highest value of rod bivalents (14.96). Quadrivalents were formed in most of the cultivars studied ranging from 0.12 (in Garison and Chinese cultivars) to 0.58 (in the cultivar Cyclone) (Table 1). Interestingly enough some cultivars possessed hexavalents ranging from 0.03 to 0.06. The residual homoeologous recombination in the cultivars studied may be the reason for quadrivalent formation in the cultivars, while hexavalents may have been formed due to subsequent chromosome translocations. ANOVA test revealed the presence of a significant difference (p<0.01) for chiasma frequency and distribution as well as bivalent and quadrivalents among the cultivars studied. Therefore at least two cultivars differ significantly on their meiotic characteristics. The LSD test showed that such significant difference is present almost among most of the cultivars, particularly those cultivars, which are, placed in different clusters/ groups in cluster analysis and ordination based on PCA (explained in the following paragraphs). No significant difference was observed for hexavalents and univalents.
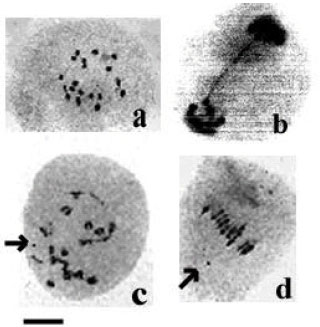 | |
| Fig.1: | a = Metaphase I cell showing 19 bivalents in the cultivar Modena, b = Anaphase I cell showing stickiness in the cultivar Cylone. c = Metaphase I cell showing B-chromosomes (arrow) in the cultivar Iris, d = Metaphase I cell showing B-chromosome (arrow) in the cultivar Modena, Scale bar = 10 μm |
| Table 1: | Meiotic characters in rapeseed cultivars studied |
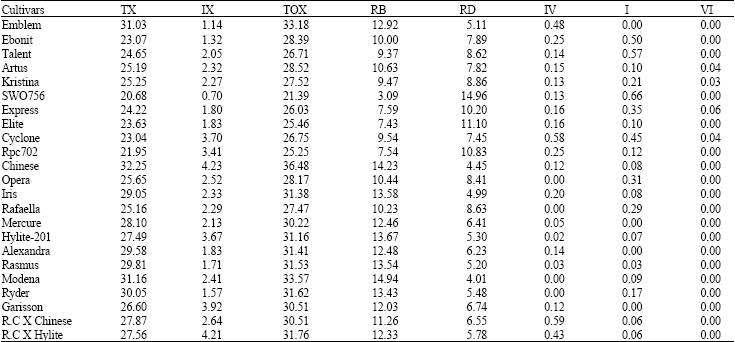 | |
TX = Terminal Chiasmata, IX = Intercalary Chiasmata, TOX = Total Chiasmata, RB = Ring Bivalent, ROB = Rod Bivalent, QU = Quadrivalent, I = Univalent, IV = Quadrivalent, VI = Hexavalent | |
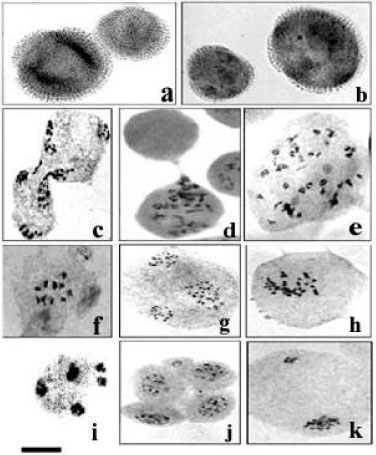 | |
| Fig. 2: | Representative meiotic cells and pollen grains in B. napus cultivars. a and b = Unreduced pollen grain (big pollen) in the cultivars Artus and Garisson, respectively. c and d = Partial and complete cytomixis in the cultivar Chinese. e = Meiocyte having double the chromosome number due to cytomixis in the cultivar Rasmus. f = Meiocyte showing reduction in chromosome number in the cultivar Chinese. g = Anaphase-II failure in the cultivar Alexandra. h = Metaphase-I cell showing Desynapsis (large number of univalents) in the cultivar Hylite-201. I = A pentapolar cell in the cultivar Elite. j = Abnormal tetrad in the cultivar Cyclone. k = Unequal segregation in the cultivar Alexandra. Scale bar = 10 μm |
The frequency and distribution of chiasma is under genetic control (Quicke, 1993), therefore presence of a significant difference in chiasma frequency and distribution as well as ring and rod bivalents among the cultivars studied may indicate partly their genomic differences as these plants were grown under uniform conditions in the experimental field.
Data with regard to chromosome segregation is provided in Table 2. Chromosome stickiness, laggard chromosomes and micronuclei occurred in most of the cultivars. The highest percentage of anaphase-I and II tickiness occurred in the cultivar Talent (14.70 and 7.27, respectively). Paired sample χ2 test showed a significant difference in the percentage of chromosome stickiness among the cultivars studied. Genetic as well as environmental factors have been considered as the reason for chromosome stickiness (De Souza and Palgliarini, 1996; Baptista-Giacomelli et al., 2000).
Anaphase-I and II laggards were observed in all the cultivars studied (Table 2). The highest value of Anaphase-I and II laggards occurred in Talent and Ebonite cultivars (14.70 and 10.00, respectively) while the lowest values occurred in Alexandra (0.39) and Elite and Express cultivars (00.00). χ2 test showed a significant difference in the percentage of laggard chromosomes among the cultivars studied indicating their genomic differences for such a meiotic behavior.
Pearson coefficient of correlation determined among meiotic irregularities themselves and also with pollen fertility revealed a positive significant correlation (p<0.05) between the percentage of laggard cells in anaphase-I with stickiness in anaphase-I and II as well as percentage of laggard cells in anaphase-II. The chromosomes stickiness in anaphase-I showed a positive significant correlation (p<0.05) with micronucleus formation. However pollen fertility did not show any correlation with these meiotic irregularities. The cultivars studied showed a high pollen fertility (> 98%, Table 2), therefore it seems that meiotic irregularities mentioned do not have a major effect on pollen fertility reduction.
| Table 2: | Meiotic abnormalities and pollen fertility in rapeseed cultivars studied |
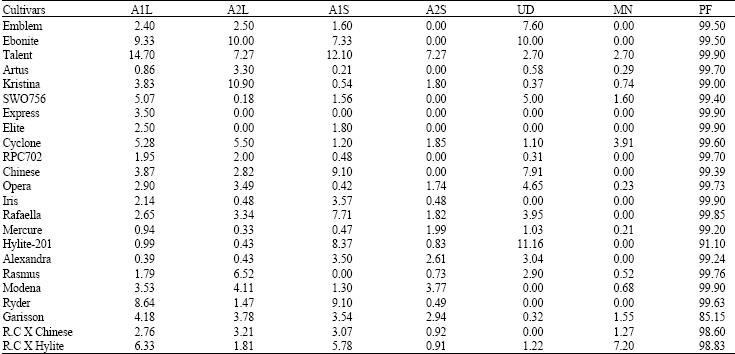 | |
A1L = Laggards in Anaphase-I, A2L = Laggards in Anaphase-II, A1S = Stickiness in Anaphase-I, A2S = Stickiness in Anaphase-II, UD = Unequal Division, MN = Micronucleus, PF = Pollen Fertility. (All values in percentage) | |
It has been suggested that infertility in polyploids is not solely due to the production of aneuploid gametes formed by improper segregation of chromosomes during anaphase/telophase stages, the genetic factors may also bring about pollen sterility as evidenced in different tetraploid strains of rye as well as Avena sativa cultivars (Baptista-Giacomelli et al., 2000). Therefore reduction in pollen fertility in B. napus cultivars may also be affected by genetic factors and not only by meiotic irregularities reported.
Different cluster analyses and ordination of cultivars based on PCA of meiotic data produced a similar result (Fig. 4-5). In general four major cluster/groups are formed, separating the cultivars studied. The first major cluster is comprised of two sub-clusters. The cultivars Opera, Rafaella, Ebonit, Talent, Elite and RPC702 form the first sub-cluster while the cultivars Artus, Kristina, Express and Cyclone comprises the second sub-cluster. Although the cultivars Express and Cyclone are joined the other cultivars of the second sub-cluster with some distance due to their meiotic differences all the four cultivars of this sub-cluster formed hexavalents in metaphase-I (Table 1). The second major cluster is also comprised of two sub-clusters. The cultivars Hylite-201, Garisson, Rasmus, Ryder, Mercure, Alexander, Iris and Modena form the first sub-cluster while the cultivars R.C X Chinese, R.C X Hylite and Emblem form the second sub-cluster. The cultivar Chinese alone forms the third major cluster and cultivar SWO756 alone forms the forth major cluster and are joined with a great distance to the other cultivars indicating their cytogenetic differences. It is interesting to mention that two hybrid cultivars of R.C x Chinese and R.C x Hyalite are obtained from hybridization of the hybrid cultivar Regen x Cobra with the cultivar Chinese and cultivar Hyalite-201 and are placed close to each other in one cluster possibly due to genomic effects of Regen x Cobra. However the parental genotypes of Hyalite-201 and Chinese are placed in different clusters. As stated earlier, the members of different clusters differ significantly in their meiotic characteristics from the others. PCA analysis of meiotic data revealed that the first 3 factors comprise about 82% of total variance. In the first factor, which comprises about 57% of total variance, meiotic characters like total and terminal chiasmata as well as ring bivalents and possessed the highest positive correlation (>0.90), while rod bivalents possessed the highest negative correlation (>-0.90). In the second factor, which comprises about 14% of total variance, quadrivalents possessed the highest positive correlation (>0.80) while in the third factor with about 10% of total variance intercalary chiasmata possessed the highest positive correlation (>0.80). Therefore these are the most variable meiotic characteristics among the cultivars studied as also revealed by ANOVA test discussed earlier. The combination of the first and second PCA factors separates mainly the members of the firs and second clusters from each other, while combination of the first and third PCA factors separates SWO756 and Chinese cultivars of the third and forth major clusters (Fig. 5a and b).
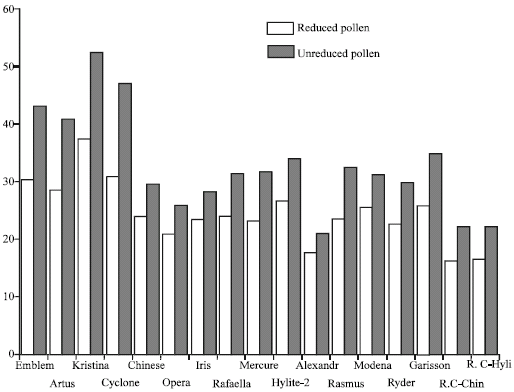 | |
| Fig. 3: | The size (μm) of reduced and unreduced pollen grains in oil rapeseed cultivars (names of the cultivars as in Table 1) |
 | |
| Fig. 4: | WARD cluster analysis of oil seed rape cultivars |
B-chromosomes: Twelve out of 23 cultivars studied showed the occurrence of 0-4 B-chromosomes (Table 3 and Fig. 1). These were smaller than the A-chromosomes and did not form any meiotic association with them, although they could arrange themselves along with the A-chromosomes on the equatorial plane of the spindle and move to the poles during anaphase. In some cases they occurred as laggard chromosomes. B-chromosomes are accessory chromosomes reported in many plant and animal species. B-Chromosomes may affect the frequency and distribution of chiasmata as well as chromosome association, either directly or by affecting the genes present on the A-chromosomes that control meiosis (Camacho et al. 2000). T-test analysis of meiotic characteristics among the cells possessing B-chromosomes compared to the cells devoid of B-chromosomes is presented in Table 3. The effect of B-chromosomes varied in different cultivars studied, for example the presence of Bs did not have any significant effect on the meiotic characters in four cultivars of Mercure, Rasmus, Garisson and R.C. X Hyalite while in the cultivars Opera, Iris, Hyalite-201 and Alexandra significantly reduced the number of intercalary chiasmata. The presence of B-chromosomes significantly increased the number of total chiasmata in the cultivars Artus, Rafaella and R.C X Chinese and significantly increased the number of intercalary chiasmata in the cultivars Artus and Ryder. A significant change in frequency of chiasmata may bring about changes in genetic recombination of the progenies this is particularly true if a significant increase in the number of intercalary chiasmata occurs.
| Table 3: | T-test analysis of meiotic characteristics among the cells possessing B-chromosomes (+ B) and cells devoid of B-chromosomes (-B) in rapeseed cultivars |
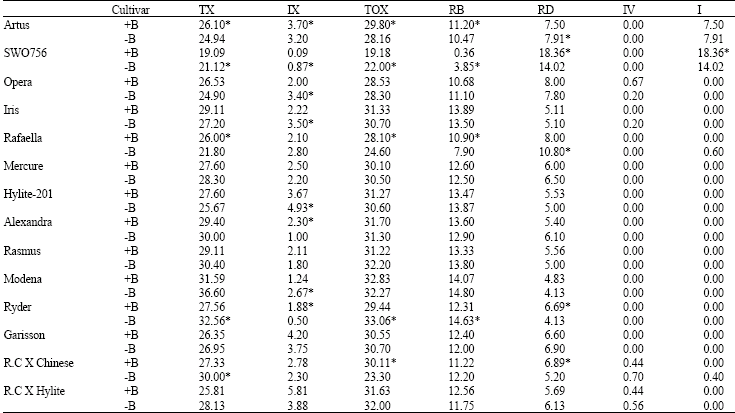 | |
Meiotic characters as in Table 1, * = Significant at p = 0.05 | |
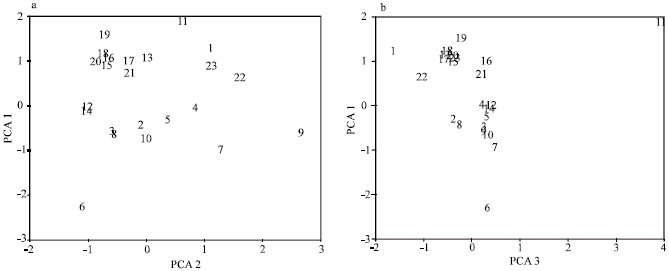 | |
| Fig. 5: | PCA ordination of oil seed rape cultivars (cultivars number as in Fig. 4) |
An increase in the number of intercalary chiasmata indicates that genes present in the middle part of the chromosomes also become involved in genetic recombination, which may lead to an increase in genetic variation of the next generation. In general if the cytogenetic differences observed in the cultivars studied is accompanied by other agronomic differences a better hybridization and selection program may be planned for B. napus cultivars.
Unreduced gamete formation: The occurrence of unreduced (2n) gametes has been considered important in the evolution of polyploids and also of economic importance in crop plants like potato for obtaining natural tetraploid plants by crossing 2x (producing unreduced gametes) X 4x lines. It was only recently that Sheidai et al. (2003) reported the occurrence of unreduced pollen grains in 6 out of 22 B. napus cultivars studied. Therefore the present study was performed to detect the occurrence of 2n pollen grains and its cytogenetic mechanisms in 23 unreported cultivars available in Iran.
The presence of meiocytes having double the gametic chromosome number as well as 2n pollen grains was noticed in 17 out of 23 B. napus cultivars studied (Fig. 3). A numerically unreduced diploid, or 2n gamete is a meiotic product that bears the sporophytic rather than the gametophytic chromosome number. Such gametes result from abnormalities during either microsprogenesis (2n pollen) or megasporogenesis (2n eggs). Unreduced gametes are known to produce individuals with higher ploidy level through a process known as sexual polyploidization (Villeux, 1985). Different methods have been used to detect 2n gametes including morphological, flow cytometery and cytological methods. The most direct method of screening for 2n pollen involves the examination of the range of size of pollens produced by an individual, as with increase in DNA content the cell volume increases which in turn influences the pollen diameter. The presence of giant pollen grains has been used as an indication of the production of 2n pollen (Vorsa and Bingham, 1979).
The measurement of the pollen grains in the B. napus cultivars with unreduced meiocytes revealed the presence of a bimodal distribution of pollen grain size. T-test analysis also showed a significant difference (p<0.01) between the pollen grains indicating the possible 2n constitution of the larger pollen grains. The frequency of 2n pollen formation varied from 0.12 in the cultivar Modena to 1.41% in the cultivar Alexandra.
In higher plants, unreduced gametes originate through two types of meiotic nuclear division: First Division Restitution (FDR) and Second Division Restitution (SDR). In case of FDR, usually failure of anaphase-I chromosomes segregation leads to the formation of restitution nucleus while in SDR failure of anaphase-II chromosomes segregation leads to the formation of unreduced gametes. However other cytological abnormalities including tripolar, parallel and fused spindles, abnormal cytokinesis as well as syncyte formation have been considered as the other causes of 2n pollen formation (Vorsa and Bingham, 1979). The present study indicates the occurrence of both FDR and SDR in B. napus cultivars and detailed cytological investigation revealed that the main cytological mechanisms for production of 2n gametes are: 1- anaphase- I and II failure in one of the poles leading to the formation of triads at the end of telophase-II instead of tetrad (one is unreduced), 2-Desynapsis or early chiasma terminalization, 3- multipolar spindles which occurred in most of the cultivars and 4- Cytomixis. Details of cytogenetical mechanisms, leading to the formation of unreduced gametes in B. napus cultivars studied are discussed bellow.
Cytomixis: Among 23 B. napus cultivars studied 13 cultivars (Table 4) showed the occurrence of cytomixis and chromosome migration (Fig. 2). The percentage of cells showing cytomixis varied from 0.79% in the cultivar Ryder to 15.76% in the cultivar Alexandra. Chromatin/ chromosome migration occurred in different directions from early prophase to telophase-II. Several metaphase/ diakinesis cells in these species possessed extra or missing chromosomes showing aneuploid condition, such aneuploid cells may lead to the formation of abnormal tetrads and infertile pollen grains. However migration of the whole chromosome complement and production of unreduced (2n) meiocytes occurred in some of the cultivars (Fig. 1), which may be the reason for production of unreduced pollen grains there after. Since the whole chromosome migration occurred before the start of anaphase I, the restitution nucleus (unreduced cell) is of FDR type. Paired wise X2 test performed for percentage of cytomixis showed a significant difference among the cultivars studied indicating their genomic difference.
Migration of chromatin material among the adjacent meiocytes occurs through cytoplasmic connections originated from the pre-existing system of plasmodesmata formed within the tissues of the anther. The plasmodesmata become completely obstructed by the deposition of callose, but in some cases they still persist during meiosis and increase in size forming conspicuous inter-meiocyte connections or cytomictic channels that permit the transfer of chromosomes (Falistocco et al., 1995). Cytomixis is considered to be of less evolutionary importance but it may lead to production of aneuploid plants or produce unreduced gametes as reported in several grass species including Dactylis and Aegilops (Falistocco et al., 1995).
| Table 4: | Unreduced pollen grain formation and the related cytogenetic mechanisms (%) of rapeseed cultivars studied |
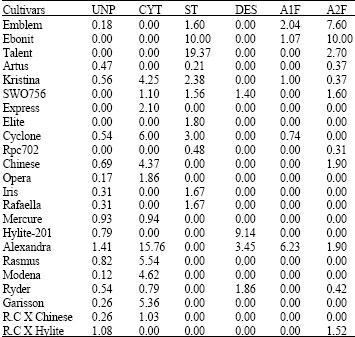 | |
UNP = Unreduced Pollen grain, CYT = Cytomixis, ST = Stickiness, DES = Desynapsis, A1F = Anaphase-I failure, A2F = Anaphase-II failure | |
Anaphase I and II failure: Failure of chromosome movement either during anaphase I or II occurred in 12 out of 23 cultivars studied. The failure of chromosome movement occurred in one of the poles of in anaphase cells (Fig. 2), leading to the formation of normal reduced and unreduced daughter cells. Such unreduced meiocytes may lead to the formation of 2n pollen grains. Since failure of chromosome segregation occurred in both anaphase-I and II, the restitution nucleus of both FDR and SDR constitution would be formed. In the cultivars Emblem, Ebonit, Kristina and Alexandra both anaphase-I and II failure occurred.
Multipolar cells: The occurrence of tripolar or multipolar cells was observed almost in most of the cultivars studied (Fig. 2). Formation of tripolar cell leads to the formation of two reduced and one unreduced pollen grains, while pentapolar cells observed in some of the cultivars may lead to the formation of abnormal tetrads and infertile pollen grains (Fig. 2). The spindle apparatus is normally bipolar and acts as a single unit, playing a crucial role in chromosome alignment during metaphase. Any distortion or breakage in the spindle may result in random sub-grouping of the chromosomes which function independently (Nirmala and Rao, 1996). In several instances spindle abnormalities have led to the production of aneuploid gametes for example in polyploidy hybrids and derivatives of Aegilopsx Triticum hybrids, amphiploid Triticineae, amphiploids of Solanum hybrids, etc. and also is considered as a reason for production of unreduced (2n) gametes in Solanum (Villeux, 1985).
Desynapsis: Desynapsis is considered as the precocious separation of bivalents in metaphase of meiosis I leading to the formation of varying degree of univalents. Partial to complete desynapsis was observed in 4 out of 23 cultivars studied (Table 4 and Fig. 2). In partial desynapsis few to some bivalents were separated to form univalents while, in complete desynapsis all bivalents were separated to form univalents. Such desynaptic cells with double the chromosome number may form unreduced pollen grains. Desynapsis occurs either due to the action of recessive ds genes in a homozygous situation or early chiasma terminalisation which may lead to the formation ofmeiocytes with double the normal chromosome number.
In several cases such univalents may have difficulty during anaphase-I movement and become lagged therefore producing aneuploid gametes causing reduction in pollen fertility of plants. However they may skip the first anaphase and form restitution nucleus resulting in the formation of unreduced gametes as reported in Solanum (Villeux, 1985).
REFERENCES
- Baptista-Giacomelli, F.R., M.S. Palgliarini and J.L. de Almeida, 2000. Meiotic behavior in several Brazilian oat cultivars (Avena sativa L.). Cytologia, 65: 371-378.
CrossRefDirect Link - Camacho, J.P.M., T.F. Sharbel and L.W. Beukeboom, 2000. B-chromosome evolution. Phil. Trans. R. Soc. Lond. B, 355: 163-178.
Direct Link - De-Souza, A.M. and M.S. Palgliarini, 1996. Spontaneous chromosome stickiness in canola. Nucleus, 39: 85-89.
Direct Link - Falistocco, E., T. Tosti and M. Falcinelli, 1995. Cytomixis in pollen mother cells of diploid Dactylis, one of the origins of 2n gametes. J. Heredity, 86: 448-453.
Direct Link - Nirmala, A. and P.N. Rao, 1996. Genesis of chromosome numerical mosaism in higher plants. Nucleus, 39: 151-175.
Direct Link - Sheidai, M., M.R. Manossorniya and M.R. Ahmadi, 2001. Cytomorphological study of some rapeseed (Brassica napus L.) cultivars in Iran. Nucleus, 44: 46-51.
Direct Link - Sheidai, M., Z. Noormohammadi and M.R. Ahmadi, 2001. Notes on B-chromosomes and SDS-PAGE proteins in some rapeseed (Brassica napus) cultivars. Nucleus, 44: 165-170.
Direct Link - Sheidai, M., M. Noormohammadi, N. Kashani and M. Ahmadi, 2003. Cytogenetic study of some rapeseed (Brassica napus L.) cultivars and their hybrids. Caryologia, 56: 387-397.
Direct Link