
Boukare Zeba
Laboratoire de Biochimie et Chimie Appliquées (LABIOCA), Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, Université de Ouagadougou 03 BP 7021 03 Ouagadougou, 03, Burkina Faso
Martin Kiendrebeogo
Laboratoire de Biochimie et Chimie Appliquées (LABIOCA), Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, Université de Ouagadougou 03 BP 7021 03 Ouagadougou, 03, Burkina Faso
Aline Lamien
Laboratoire de Biochimie et Chimie Appliquées (LABIOCA), Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, Université de Ouagadougou 03 BP 7021 03 Ouagadougou, 03, Burkina Faso
Jean-Denis Docquier
CIP/Enzymology (Université de Liège) Institute of Chemistry B6 Sart-Tilman B4000 Liège, Belgium
Jacques Simpore
Laboratoire de Biochimie et Chimie Appliquées (LABIOCA), Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, Université de Ouagadougou 03 BP 7021 03 Ouagadougou, 03, Burkina Faso
Odile Germaine Nacoulma
Laboratoire de Biochimie et Chimie Appliquées (LABIOCA), Unité de Formation et de Recherche en Sciences de la Vie et de la Terre, Université de Ouagadougou 03 BP 7021 03 Ouagadougou, 03, Burkina Faso
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 3 | Page No.: 506-510







ABSTRACT
Many clinical species of bacteria were isolated from biological samples such as urines, blood and wound in Saint Camille medical centre of Ouagadougou. Among the concerned species, the most important members were Escherichia coli and Klebsiella pneumoniae. These β-lactamases producing isolates were directly screened by PCR to identify the nature of the amplified genes responsible for penicillin destroying activity. Therefore specific TEM and SHV primers were used. The PCR products were sequenced. The sequencing results indicated that the parental forms blaTEM-1 and blaSHV-1 were the most common determinants of β-lactamase found, respectively in Escherichia species and Klebsiella pneumoniae. The bacterial susceptibility analysis by MICs measurement clearly correlated the presence of concerned β-lactamase determinants and their resistance patterns. This study is part of a set of investigations carried out by our laboratory to assess the β-lactamase incidence in the failure of β-lactam therapy. In particular, the purpose of this study was to determine the precise nature of β-lactamase supporting the low susceptibility of host bacteria towards penicillins.
PDF Abstract XML References Citation
How to cite this article
Boukare Zeba, Martin Kiendrebeogo, Aline Lamien, Jean-Denis Docquier, Jacques Simpore and Odile Germaine Nacoulma, 2007. Major Enzymatic Factors Involved in Bacterial Penicillin Resistance in Burkina Faso. Pakistan Journal of Biological Sciences, 10: 506-510.
DOI: 10.3923/pjbs.2007.506.510
URL: https://scialert.net/abstract/?doi=pjbs.2007.506.510
DOI: 10.3923/pjbs.2007.506.510
URL: https://scialert.net/abstract/?doi=pjbs.2007.506.510
INTRODUCTION
Antibiotics are widely used to fight pathogenic bacteria. Penicillin was the first antibiotic discovered seventy-eight years ago (Fleming, 1929). Penicillin belongs to a vast family of molecules including cephalosporins, carbapenems and monobactams. All these compounds bear the β-lactam ring which is their reactive centre. The appearance of enzymes from bacteria (Abraham and Chain, 1940) able to destroy penicillin and related molecules was the beginning of a severe war between pathogenic bacterial cells and man. These penicillin inactivating enzymes known as β-lactamases secreted by bacteria were subdivided into four classes A, B, C and D on the basis of sequence characteristics (Ambler, 1975, 1980; Ambler and Scott, 1978).The class A enzymes are numerically the most important group of β-lactamases found in many clinical isolates of Enterobacteriaceae worldwide. These enzymes are often inhibited by suicide inactivators such as clavulanic acid (Bush, 1988) which is clinically used to protect β-lactamase susceptible antibiotics. This class comprises two main representatives named TEM and SHV. The term TEM comes from Temoniera, the name of the patient in Greece from which the host bacteria of the first enzyme of this species was isolated (Medeiros, 1984). TEM-1 is the parental form. The word SHV comes from Sulphydryl Variable the biochemical properties (Barthelemy et al., 1986) of this species whose parental form is SHV-1. TEM-1 and SHV-1 share 68% homology in amino-acid sequence (Du Bois et al., 1975; Majiduddin et al., 2003). The two β-lactamases have also similar substrate profiles in that both can readily hydrolyse β-lactams belonging to penicillin group such as benzylpenicillin, ampicillin, amoxicillin piperacillin etc. However, it was reported that SHV-1 achieves better activity against ampicillin (Bush et al., 1995).
Resistance to β-lactam antibiotics has been a problem for as long as these drugs have been used in clinical practice. Throughout the world, the problem of resistance to β-lactam antibiotic via the production of β-lactamase has become a serious public health preoccupation. Until now, no significant information on the nature of β-lactamase involved in the clinical isolates resistance to penicillin in Burkina Faso (West Africa) was available. This study was a real attempt to fulfil this gap in order (1) to characterize the type of Escherichia coli and Klebsiella pneumoniae β-lactamase directly being involved in penicillin resistance in Burkina Faso (2) to eventually highlight derivatives of existent enzymes.
MATERIALS AND METHODS
Bacterial isolates: The isolates analysed in this study were all β-lactamase producing enterobacteriaceae collected at the Saint Camille Medical Centre (SCMC), a central institution for care and medical analysis in the capital city-Ouagadougou. The sources of the isolates were mainly faeces, urines, pu etc. They were identified by Api 20 and Vitek 2 Systems (BioMerieux France). One hundred organisms were selected solely because of the substrate profile of their β-lactamase and their resistance to penicillins as reported by the laboratory analysis. Thirteen representative isolates were retained for further investigation. They include: 2 Escherichia fergusonii (N°672 and 681) 5 Escherichia coli (N°685, 1004, 1181, 1190, 1204 1220) and 5 Klebsiella pneumoniae (N°Z54, 291, 312, 392, 1201).
Antibiotic susceptibility: Bacterial species analysed were phenotypically characterized by determining their susceptibility towards different penicillins. Therefore, the Minimum Inhibitory Concentrations (MICs) were determined using the automatic method VITEK 2 (BioMerieux In., St Louis, Mo.)
Molecular study: Considering that Escherichia species and Klebsiella pneumoniae often carry TEM or SHV-β-lactamase, the genes encoding these enzymes were targeted by PCR amplification directly on bacterial cells using TEM and SHV specific primers. The different Primers used for amplification are shown in Table 1. These oligonucleotides were obtained from Eurogentec Bel. S.A. Cycling conditions were as follow for blaSHV genes amplification: 24 cycles of denaturation at 94°C for 1 mn, annealing at 37°C for 1 mn and extension of primers at 72°C for 1 mn. Cycling conditions for blaTEM amplification were the same except that anneling temperature was raised to 45°C. All reactions were carried out in a final volume of 50 μL containing 1U of Taq or GoTaq DNA polymerase, 8 pM of Each primer, 200 μM of deoxynucleotide triphosphate and 2 μL of crude bacterial lysate (obtained by boiling bacterial suspensions in distilled water for 20 mn).
| Table 1: | Oligonucleotides used as primers for amplification of blaSHV and blaTEM genes |
 | |
This crude bacterial lysate was used as template. PCR amplicons were immediately purified by Wizard purification kit of PCR products.
RESULTS
Antibiotic susceptibility: All the clinical isolates analysed were β-lactamase-positive and the corresponding enzymes were active on benzylpenicillin, ampicillin, amoxicillin, carbenicillin, oxacillin and piperacillin. The results obtained from the analysis of their susceptibility towards these β-lactams and some combinations are reported in Table 2.
PCR results: A total of 13 penicillin resistant and β-lactamase producing isolates including Escherichia species and Klebsiella pneumoniae were screened by PCR to elucidate the determinants of enzymes involved in major penicillins (benzylpenicillin, ampicillin, amoxicillin) resistance. From the 13 isolates, 3 Klebsiella pneumoniae (N°291, 312 and 392) were analysed in another experiment (results not shown). The PCR amplification was successful with all the isolates screened as shown in Fig. 1. The amplicons generated were good in term of quantity and quality. The fragments amplified using TEM and SHV primers were all about 800 base pairs (Fig. 1). These sizes correspond approximately to those of TEM and SHV nucleotide sequences. Genes encoding TEM-types were generated from Escherichia coli, while SHV genes were obtained from Klebsiella pneumoniae isolates. Klebsiella pneumoniae isolate N°1201 was found to carry both TEM and SHV types of genes.
Sequence analysis: All the amplicons obtained were sequenced after purification. As shown from Fig. 1, isolate N°1201 appeared twice because of carrying simultaneously SHV and TEM genes. The complete nucleotide sequences of most of the fragments were obtained (Table 3) except 2 amplicons where the whole sequence could not be determined. These amplicons were from Klebsiella pneumoniae N°291 and Escherichia coli N°1004. But amplicon N°291 was identified to be a SHV determinant and amplicon 1004 was a TEM one. The lack of full sequence did not enable the exact assignment of the determinant type (parental or variant in the two cases).
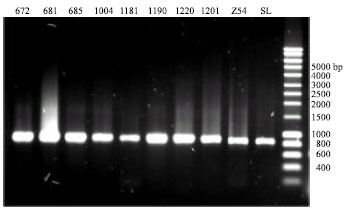 | |
| Fig. 1: | Analysis of purified PCR fragments from cell lysate by agarose 1% gel electrophoresis amplicons N°672 and 681 were obtained from Escherichia fergusonii. Amplicons N° 685, 1004, 1181, 1190, 1201, 1220 were obtained from Escherichia coli. amplicons N°1201 and Z54 were generated from Klebsiella pneumoniae. Two fragments bear the same number (1201) on the figure because of the generation of 2 amplicons from SHV and TEM primers using the same template (lysate from Klebsiella pneumoniae N°1201). Smart Ladder (SL) is the DNA marker. The size of the markers in base pairs (bp) is shown on the right |
DISCUSSION
Various isolates (Escherichia species and Klebsiella pneumoniae) from human biological samples at Saint Camille Medical Centre of Ouagadougou and supposed to be responsible for infections (urinary, intra abdominal disease, wound etc.) were analysed. Their potency to resist to penicillin was investigated and correlated to their contents of β-lactamase determinants. Most the isolates were found to have low susceptibility towards the three major penicillins as shown by the values of their MICs (Table 2). Isolate N°1204 was quite different from the others and appeared to be a negative control owing to its overall susceptibility to penicillin molecules. This behaviour could suggest the very low level of the β-lactamase expression in this isolate. Sequencing of blaSHV and blaTEM genes isolated by PCR amplification from the corresponding isolates identified the exact type of β-lactamase determinant. All the Escherichia species (coli and fergusonii) carried blTEM-1,a while most of the Klebsiella pneumoniae carried blaSHV-1. Nevertheless one Klebsiella pneumoniae was found to carry the derivative blaSHV-11 while another appeared to carry both blaSHV-11 and blaTEM-1.
| Table 2: | MICs of β-lactams for clinical isolates analysed |
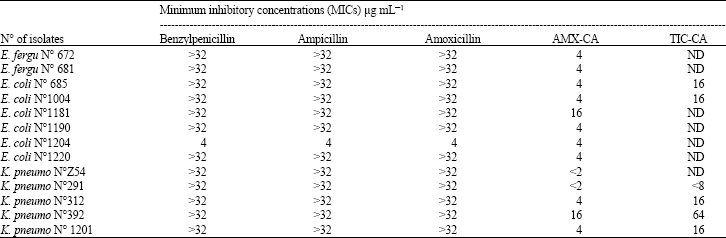 | |
Abbreviations used in Table 2, AMX-CA = Amoxicillin-clavulanic acid, TIC-CA = Ticarcillin-clavulanic acid, E. coli = Escherichia coli, E. fergu = Escherichia fergusonii, K. pneumo = Klebsiella pneumoniae, ND = Not Determinated | |
| Table 3: | Types of β-lactamase encoding genes |
 | |
| Table 3: | Continued |
 | |
Normally the presence of two determinants would confer an exceptional high level of resistance to this isolate. Indeed, it was quite resistant to the three penicillins reported above but the profile was not exceptional probably because of low constitutive expression level of the two genes. About the isolates N°291 and 1004 where detection of SHV and TEM gene was obtained respectively but no full sequences, it is improbable for these organisms to carry other determinants than blaSHV-1 or blaTEM-1 owing to the substrate profiles of their extracts and their susceptibility patterns. It should be noted that there has been few data about β-lactamase investigation in Africa, except South Africa where substantial data on the subject were available. So blaSHV-1 and many derivatives (-2, -5, -19, -20, -21 and -22) and blaTEM-1 with some derivatives (-53, -63) were reported in South Africa (Pitout et al., 1998; Essack et al., 2001). South African data include an important number of Extended Spectral β-Lactamase (ESBL) determinants among blaSHV genes as well as blaTEM genes isolated from Klebsiella pneumoniae and Escherichia coli species. The list of African countries where ESBL are emerging grows with a recent report from Cameroon/Central Africa (Gangoué-Pieboji et al., 2005). However, the current data obtained in Burkina Faso do not show the presence of ESBL into the two types of β-lactamase encoding genes. The results obtained from the screened isolates strongly suggest the occurrence of SHV-1 and TEM-1 enzymes and their probable role in penicillins resistance. These conclusions are consistent with evidence that SHV-1 and TEM-1 enzymes generally demonstrate greater turnover numbers (kcat) for benzylpenicillin, ampicillin and amoxicillin (Hujer et al., 2001; Maveyraud et al., 1996; Dubois et al., 2004). Hitherto no ESBL (efficient derivative from SHV-1 or TEM-1 enzyme) has been identified in our investigations. But this does not mean that ESBL are absent in our country. In the local antibiotherapy background a rare enzyme of high clinical relevance and multiple β-lactam resistance factor such as metallo-β-lactamase was reported in Burkina Faso (Zeba et al., 2005). Before this evidence, the hypothesis was that the antibiotherapy conditions were not favourable for the emergence of this type of enzyme. This means that ESBL existence is not impossible. However, the small number of isolates analysed could also explain why none of ESBL was not encountered yet. Investigations are underway to establish the possible ESBL existence and the extent of their involvement in bacterial drug resistance.
In summary, this study shows the existence of SHV-1 and its variant SHV-11 that were carried by Klebsiella pneumoniae in Burkina Faso. Likewise, TEM-1 was found in Escherichia species. SHV-1, its derivative SHV-11 and TEM-1 are probably the major enzymatic factors involved in penicillins resistance of the concerned species.
ACKNOWLEDGMENTS
This study was supported by the Coopération Universitaire au Développement (CUD) from Belgium Kingdom through a project named Etude des enzymes de la résistance bactérienne aux antibiotiques. We greatly thank this institution. We also thank Professor Jean-Marie Frère Head of CIP (Protein engineering Center) in University of Liège/Belgium where the molecular study was carried out. A great thank was also addressed to Dr. Colette Duez and Pierrette Melin for their useful technical contribution.
REFERENCES
- Abraham, E.P. and E. Chain, 1940. An enzyme from bacteria able to destroy penicillin. Nature, 146: 837-837.
CrossRefDirect Link - Ambler, R.P., 1980. The structure of β-lactamases. Philos. Trans. R. Soc. Lond. B: Biol. Sci., 289: 321-331.
PubMedDirect Link - Bush, K., 1988. β-lactamase inhibitors from laboratory to clinic. Clin. Antimicrob. Chemother., 1: 109-123.
CrossRefDirect Link - Bush, K., G.A. Jacoby and A.A. Medeiros, 1995. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother., 39: 1211-1233.
PubMedDirect Link - Dubois, V., L. Poirel Carping, L. Coulange, C. Bebear, P. Nordmann and C. Quentin, 2004. SHV-49, a novel inhibitory resistant β-lactamase in clinical isolate of Klebsiella pneumonie. Antimicrob. Agents Chemother., 48: 4466-4469.
Direct Link - Essack, S.Y., L.M.C. Hall, D.G. Pillay, M.L. McFadyen and D. Livermore, 2001. Complexity and diversity of Klebsiella pneumoniae Strains with Extended-Spectrum β-lactamases isolated in 1994 and 1996 at a Teaching Hospital in Durban, South Africa. Antimicrob. Agents Chemother., 45: 88-95.
CrossRef - Majiduddin, F.K. and T. Palzkill, 2003. An analyse of why highly similar enzymes evolve differently. Genetics, 163: 456-466.
Direct Link - Pitout, J.D.D., K.S. Thompson, N.D. Hanson, A.F. Ehrhardt, E.S. Moland and C.C. Sander, 1998. β-lactamases responsible for resistance to expanded-spectrum cephalosporins in Klebsiella pneumoniae, Escherichia coli and Proteus mirabilis isolates recovered in South Africa. Antimicrob. Agents Chemother., 42: 1350-1354.
Direct Link - Zeba, B., J. Simpore, O. Nacoulma and J-M. Frere, 2005. Identification of metallo-β-lactamase from a clinical isolate at saint camille medical center of ouagadoudou, Burkina Faso. Afr. J. Biotechnol., 4: 286-288.
Direct Link - Du Bois, S.K., M.S. Marriott and S.G. Amyes, 1975. TEM- and SHV-derived extended-spectrum β-lactamases: Relationship between selection, structure and function. J. Antimicrob. Chemother., 35: 7-22.
PubMedDirect Link