
F. Hoori
Department of Biology, Faculty of Science, Isfahan University, Isfahan, Iran
A.A. Ehsanpour
Department of Biology, Faculty of Science, Isfahan University, Isfahan, Iran
A. Mostajeran
Department of Biology, Faculty of Science, Isfahan University, Isfahan, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 3 | Page No.: 481-485







ABSTRACT
In this study, the regeneration through embryogenesis of two species of Medicago were studied. Seeds of Medicago sativa cv. Rehnani and M. truncatula line A17 were grown on MS medium. After 4-6 weeks, segments of leaf and stem from two species were transferred to MS medium containing 2 mg L-1 NAA, 2,4-D and Kinetin. The results indicated that callus formation from leaf explants of M. sativa was higher than M. trancatula. In the next stage, media with different combinations of auxin, cytokinin or ethinyl estradiol were provided for regeneration. Then in two stages, explants of leaf and stem of two species were transferred on these media. Results after 3-6 weeks showed that in medium containing NAA and TDZ, stem pieces of M. sativa produced shoots while leaf pieces on NAA and ethinyl estradiol formed roots. Leaf explants of M. truncatula in the medium containing NAA and BAP, produced somatic embryos. Also in media with auxin and ethinyl estradiol, somatic embryos were formed on calli of two species. Ethinyl estradiol and auxin together can induce somatic embryogenesis and root production on calli and stem or leaf explants.
PDF Abstract XML References Citation
How to cite this article
F. Hoori, A.A. Ehsanpour and A. Mostajeran, 2007. Comparison of Somatic Embryogenesis in Medicago sativa and Medicago truncatula . Pakistan Journal of Biological Sciences, 10: 481-485.
DOI: 10.3923/pjbs.2007.481.485
URL: https://scialert.net/abstract/?doi=pjbs.2007.481.485
DOI: 10.3923/pjbs.2007.481.485
URL: https://scialert.net/abstract/?doi=pjbs.2007.481.485
INTRODUCTION
Medicago truncatula Gaertn. (known as barrel medic because of the shape of its seeds pods) is an annual species (Fabaceae) that is native to the Mediterranean area and is widely grown as a pasture legume and crop rotation in a number of regions throughout the world (Crawford et al., 1989). This species is an autogamous diploid species (2n = 2X = 16) with a small genome (Neves et al., 1999). This plant species is self fertile and has short life cycle of about 3 months (Barker et al., 1990; Araujo et al., 2004). These characteristics enables this species to be used in molecular genetic studies like analysis of gene expression, promoter functional analysis, T-DNA mutagenesis and expression of genes for crop improvement (Somers et al., 2003). It is also a model plant for the study of legume-rhizobium symbiosis (Barker et al., 1990).
Medicago sativa L. is not annual species but is closely related to M. truncatula. This species has high degree of heterozygosity and large genome size (2n = 32) (Iantcheva et al., 1999).
Legumes have been regarded as recalcitrant to transformation and their in vitro regeneration is highly genotype dependent and only rarely cultivated varieties are amenable to regeneration (Somers et al., 2003).
Regeneration through somatic embryogenesis is frequently of single cell origin, resulting in a low frequency of chimera and a high number of regenerants (Neves et al., 1999). These characteristics render somatic embryogenesis an attractive system for the introduction of gnomic traits of interest by genetic engineering (Neves et al., 1999). Usually an auxin is required to induce somatic embryogenesis and subsequent auxin withdrawal or lowering of the auxin concentration is required for embryo maturation (Dudtis et al., 1991; Nolan et al., 2003; Von Arnold et al., 2002). Auxin play critical roles in the major growth responses during plant development. At cellular level, auxin acts as a signal for division, expansion and differentiation throughout the plant life cycle. By contrast less attention has been directed to effect of steroids on plants. In addition to the hormone regime, key variables are explant, developmental stage, nutrition regime and genotype (Rose and Nolan, 1995). Regeneration via organogenesis or embryogenesis are the basis of tissue culture methods and without regeneration, it is impossible to produce transgenic plants.
In this study, we considered the effect of several growth regulators including ethinyl estradiol on somatic embryogenesis of M. truncatula line A17 and M. sativa cv. Rehnani.
MATERIALS AND METHODS
Seeds of M. truncatula line A17 were obtained from State Agriculture and Biotechnology Center ( SABC) WA. Seeds of M. sativa cv. Rehnani were supplied from Seed and Seedling Research Center of Isfahan.
| Table 1: | MS medium with plant growth regulators for plant regeneration |
 | |
Seeds of M. truncatula first scarified by sands for germination. Then seeds of both species were surface sterilized for 30 sec with ethanol (70% v/v) followed by 30% (v/v) sodium hypochlorite for 15 min. After 3-4 rinses by sterile distilled water, seeds of M. sativa were cultured on MS medium (Murashige and Skoog, 1962) supplemented with 3% (w/v) sucrose and solidified with 1% (w/v) agar while, seeds of M. truncatula first were cultured on 1% (w/v) water agar medium and after two weeks seedlings were transferred to MS medium with the same combination as described already. The pH of all media was adjusted to 5.8 before autoclaving (15 min at 121°C).
For callus induction, leaf and stem segments of in vitro grown seedlings were transferred to callus proliferation, C medium.
Cultures were maintained in the culture room with 16/8 light-dark photoperiod at 25±2°C. The experiments were carried out with 10 replications and 4 explants in each replication. In the next stage, after two subcultures calli were transferred to MS medium supplemented with different growth regulators according to Table 1.
Embryogenic calli then were transferred to hormone free medium to develop somatic embryos.
All data were analyzed according to Duncan or Chi-Square tests using SPSS and Sigma Stat programs.
RESULTS
Stem and leaf explants of M. sativa and M. truncatula on C medium produced high percentage of soft calli as Table 2 showing fresh weight of callus resulted from stem is relatively lower than leaf.
When calli obtained from either stem or leaf were transferred to 7 different media for somatic embryogenesis, a few media induced somatic embryos in both Medicago species. For instance, leaf and stem callus produced the highest percentage of embryos in R1 medium (Table 3), while, in Medicago truncatula leaf and stem callus produced the highest percentage of somatic embryo in R3 and R1 media, respectively (Table 4). In other media with different combination of plant growth regulators we observed different percentage of somatic embryo initiation within 20-30 days after calli were transferred to the embryogenesis media.
| Table 2: | Callus production in C medium. Uncommon letters are significant (p<0.05) |
 | |
| Table 3: | The effects of different combination of growth regulators on somatic embryogenesis of Medicago sativa callus |
 | |
| Table 4: | The effects of different combination of growth regulators on somatic embryogenesis of Medicago truncatula callus |
 | |
| Table 5: | Comparing the number of somatic embryos formed per callus of stems and leaves in M. sativa and M. truncatula in different media based on Chi-Square analysis on 5% (comparisons are columnar). Uncommon letters are significant (p<0.05) |
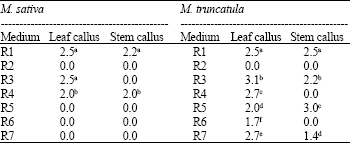 | |
| Uncommon letter (s) are significant | |
Development of somatic embryos from leaf segments of M. sativa and M. truncatula are illustrated in Fig. 1.
In R5 medium, calli obtained from stems and leaves of M. sativa grew very well and became green and after 35 days formed shoot buds.
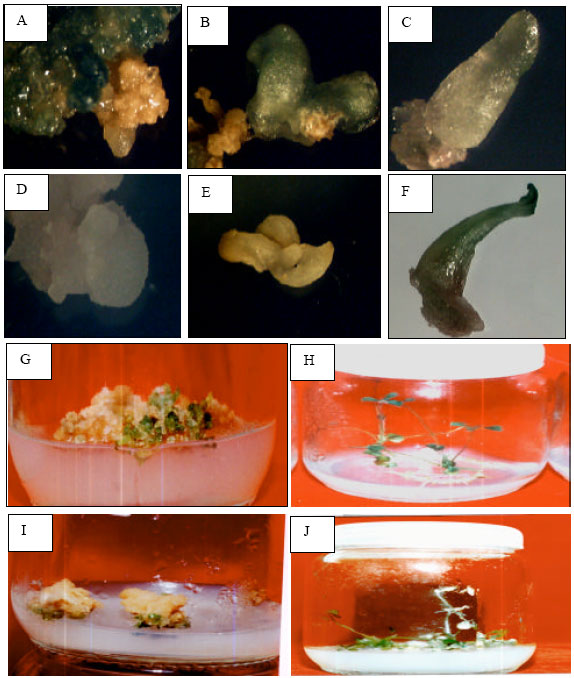 | |
| Fig. 1: | Different stages of somatic embryo development From M. sativa (A, B, C, G, H), M. truncatula (D, E, F, I, J), letters are: globular stage, heart stage, torpedo stage, plant let formation, mature plant, respectively |
In this medium, leaf segments produced green calli but no shoots were developed. Comparing of somatic embryogenesis data for both Medicago species in different media are illustrated in Table 5.
DISCUSSION
We present here an efficient procedure for somatic embryogenesis using a suitable combination of plant growth regulators in particular using ethanol estradiol for Medicago trucatula line A17 and Medicago sativa cv. Rehnani. At the first step, calli were formed on MS medium containing 2 mg L-1 of each three growth regulators (NAA, 2,4-D and kinetin). In respect to R7 medium (NAA 0.5 mg L-1), we concluded that NAA alone is not enough for callus formation but combination of auxin and cytokinin and their possible interaction in the medium promote callus formation. Our results agree with Saunders and Bingham (1972). They reported that B-II basal medium supplemented with 2 mg L-1 of NAA, 2,4-D and kinetin produced calli from anther, cotyledons, hypocotyls and internodes culture of M. sativa within 4 weeks. Nolan and Rose (1998) has also showed that NAA alone can not induce callus formation in M. truncatula. It has been well documented that 2,4-D (a strong growth regulator) with combination of cytokinin and yeast extract as a source of organic nitrate stimulates callus formation and somatic embryogenesis in M. sativa (McKersie and Brown, 1997, Saunders and Bingham, 1972). We found that this combination of plant hormones in MS medium promote callus formation from M. truncatula.
The responses of various explants to the medium is depending to the type of explant. For example, leaves were more responsive explants for dedifferentiation and callus formation in Medicago sativa while, stems produced more callus than leaves in M. truncatula.
Our data showed that NAA or ethinyl estradiol alone could not form callus but rather induced root formation. Estradiol is the strongest form of natural estrogen that resembles in central nucleus to cholesterol and is synthesized from testosterone (Rawn, 1989). Ethinyl estradiol may have auxin-like activity that its combination with NAA strengthened its effect and cause root formation from leaf explants of M. sativa.
In R2 medium, 4.2% of leaf explants of M. truncatula developed plantlets via indirect regeneration. It has been shown that NAA and BAP in the medium are very effective for induction of somatic embryogenesis of M. truncatula (Nolan and Rose, 1998; Nolan et al., 1989).
McKersie and Brown (1997) reported that IAA is not an effective auxin in callus formation and induction of somatic embryogenesis in M. sativa. Application of ethinyl estradiol alone (R6 medium) had no effects on embryo development. Therefore, combination of IAA and ethinyl estradiol (R3) had better effect on embryo formation.
Explants of leaves and stems of M. sativa in R5 medium formed calli but only stem pieces produced shoots. The auxin/cytokinin ratio is an important factor for regeneration. Increasing of cytokinin level may lead to shoot formation. Our results indicated that in R1 medium root and embryogenic masses formation may occur from interaction of auxin and ethinyl estradiol in this medium. According to some datas, auxin acts synergistically with estradiol (Goda et al., 2004; Bajguz and Tretyn, 2003). Cytokinin-like activity of ethinyl estradiol in combination with auxin might be another reason for embryo formation (Bao et al., 2004).
TDZ is a synthetic compound which is a derivative of phenylurea and has been reported to possess strong cytokinin-like activity in a number of plants (Huetteman and Preece, 1993). It has been used successfully for shoot regeneration in diverse plant species including woody plants and leguminous plants such as beans and peanut (Matand et al., 1994). However, presence of TDZ probably prevented somatic embryogenesis in calli derived from leaves and stems of M. sativa and M. trancatula.
Our data are in contrast with Iantcheva et al. (1999), they proposed that high frequency of somatic embryogenesis induced by TDZ might influence the endogenous level of cytokinins, auxins and abscisic acid so as induce the positive embryogenic response of cultivated tissue.
It can be concluded that embryogenic potential of many Medicago species is genotype specific. In addition, type of explant, its genetic potential, combination of plant growth regulators in the medium and endogenous level of hormone in tissues are the most effective factors on somatic embryogenesis and plant regeneration.
ACKNOWLEDGMENT
Authors thank the Graduate Directorate of Isfahan University for their support of this project.
REFERENCES
- Bajguz, A. and A. Tretyn, 2003. The chemical characteristic and distribution of brassinosteroids in plants. Phytochemistry, 62: 1027-1046.
Direct Link - Araujo, S., A. Duque, D. Santos and M. Fevereiro, 2004. An efficient transformation method to regenerate a high number of transgenic plants using a new embryogenic line of Medicago truncatula cv. Jemalog. Plant Cell Tissue Organ Cult., 78: 123-131.
Direct Link - Bao, F., J. Shen, S. Brady, G. Muday and T. Asami, 2004. Brassinosteroids intereract with auxin to promote lateral root development in Arabidopsis. Plant Physiol., 134: 1624-1631.
CrossRefDirect Link - Barker, D.J., S. Bianchi, F. Blondon, Y. Dattee and G. Duc et al., 1990. Medicago truncatula, a model plant for studying the molecular genetics of the Rhizobium-legume symbiosis. Plant Mol. Biol. Rep., 8: 40-49.
CrossRefDirect Link - Goda, H., S. Sawa, T. Asami, S. Fujioka, Y. Shimada and S. Yoshida, 2004. Comprehensive comparison of auxin-regulated genes in Arabidopsis. Plant Physiol., 134: 1555-1573.
Direct Link - Huetteman, C.A. and J.E. Preece, 1993. Thidiazuron: A potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult., 33: 105-119.
CrossRefDirect Link - Iantcheva, A., M. Vlahova, E. Balakova, E. Kondorosi, M.C. Elliott and A. Atanassov, 1999. Regeneration of diploid annual medics via direct somatic embryogenesis promoted by thidiazuron and benzylaminopurine. Plant Cell Rep., 18: 904-910.
CrossRefDirect Link - Matand, K., D. Porodo and C.S. Prakash, 1994. Thidiazuron promotes high frequency regeneration of peanut (Arachys hypogaea) plants in vitro. Plant Cell. Rep., 14: 1-5.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nolan, K.E., R.R. Irwanto and R.J. Rose, 2003. Auxin up-regulates MtSERK1 expression in both Medicago truncatula root forming and embryogenic cultures. Plant Physiol., 133: 218-230.
Direct Link - Von Arnold, S., I. Sabala, P. Bozhkov, J. Dyachok and L. Filonova, 2002. Developmental pathways of somatic embryogenesis. Plant Cell. Tissue Organ Cult., 69: 233-249.
CrossRefDirect Link - Somers, D.A., D.A. Samac and P.M. Olhoft, 2003. Recent advances in legume transformation. Plant Physiol., 131: 892-899.
Direct Link