
K. Nofouzi
Department of Pathobiology, School of Veterinary Medicine, Shahid Chamran University, Ahwaz, Iran
M.R. Seyfi Abad Shapouri
Department of Pathobiology, School of Veterinary Medicine, Shahid Chamran University, Ahwaz, Iran
M. Jamshidian
Department of Pathobiology, School of Veterinary Medicine, Shahid Chamran University, Ahwaz, Iran
M. Mayahi
Department of Clinical Sciences, School of Veterinary Medicine, Shahid Chamran University, Ahwaz, Iran
M. Ghaforian
Department of Immunology, Medical College, Jundi Shapour Medical Sciences University, Ahwaz, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 3 | Page No.: 470-473







ABSTRACT
Ornithobacterium rhinotracheale (ORT) infections cause major losses to the poultry industry. In search for factors implicated in the pathogenesis of ORT infections, the role of outer membrane proteins (OMPs) in the interaction of ORT with chicken tracheal epithelium was investigated. For this purpose, immune sera were prepared against total extracted OMPs, whole cell bacteria and three major OMPs of 45, 53 and 70 kDa and used in bacterial adherence inhibition assay. The results showed antibodies against a 53 kDa OMP significantly (p<=0.05) inhibited the bacterial adherence to chicken tracheal epithelium up to 78%.
PDF Abstract XML References Citation
How to cite this article
K. Nofouzi, M.R. Seyfi Abad Shapouri, M. Jamshidian, M. Mayahi and M. Ghaforian, 2007. The Role of Outer Membrane Proteins of Ornithobacterium rhinotracheale
in Attachment to Chicken Tracheal Epithelium. Pakistan Journal of Biological Sciences, 10: 470-473.
DOI: 10.3923/pjbs.2007.470.473
URL: https://scialert.net/abstract/?doi=pjbs.2007.470.473
DOI: 10.3923/pjbs.2007.470.473
URL: https://scialert.net/abstract/?doi=pjbs.2007.470.473
INTRODUCTION
Ornithobacterium rhinotracheale (ORT) is a pleomorphic, rod-shaped, gram-negative bacterium associated with respiratory disease in poultry. The poultry industry has suffered significant financial losses because of the drop in egg production, growth suppression, mortality and condemnation of carcasses in flocks infected with this organism (Van Empel and Hafez, 1999; Lopes et al., 2000; Hafez, 2002; Chansiripornchai, 2004). The bacterium primarily infects the trachea, lungs and air sacs, but can also manifest as a systemic disease with hepatitis, joint lesions and cerebrovascular pathology (Van Empel et al., 1997; Van den Bosch, 1998; Schuijffel et al., 2005). Concomitant infections with other respiratory pathogens appear to aggravate the symptoms but are not required for causing disease (Schuijffel et al., 2005). At this time, up to 18 different serotypes of ORT have been identified (Van den Bosch., 1998; Schuijffel et al., 2005). Serotype A represents 94% of all isolates from chickens, whereas it represents 57% of isolates from turkeys. Serotype D and F have not been isolated from chickens, nor has Serotype G been isolated from turkey (Joubert et al., 1999).
O. rhinotracheale control can not be achieved effectively through antibiotic use. Results from antibiotic sensitivity tests of 45 strains, collected at 45 different broiler farms with clinical respiratory disease, has demonstrated that acquired resistance against commonly used antibiotics is very high (Cauwerts et al., 2002).
The mechanisms via which ORT causes disease are largely unknown. Pathology on infected animals suggests that the bacteria colonize the respiratory tract of the host by adhering to host mucosa. Treatment of the disease is complicated, as most isolates of ORT are resistant to most commonly used antibiotics. Vaccination is an effective form of infection prevention. Available vaccines composed of killed ORT however, are assumed to give only partial protection against a limited number of strains (Van den Bosch, 1998).
To understand the role of outer membrane proteins (OMPs) in the pathogenesis of ORT and also to contribute in developing novel strategies for immunization of chickens against the bacteria, we studied the effect of polyclonal anti-OMPs sera in bacterial adherence inhibition assays.
MATERIALS AND METHODS
Bacterial cultures: An ORT serotype A, isolated from commercial chickens was used. Bacteria were grown onto Columbia agar (Lab M, UK) for 48 h at 37°C in 5% CO2. Bacterial colonies were collected, washed three times with phosphate-buffered Saline (PBS), pH 7.4 and finally resuspended in PBS to yield an optical density of 0.6 at 550 nm. The suspension contained 3-3.5x108 colony-forming units of ORT per mL and was used for immunization and adherence studies.
Extraction of OMPs: For extraction of OMPs, ORT was first grown (48 h, 37°C, 5% CO2) onto Columbia agar and then in 200 mL of Todd-Hewitt broth (Biomark, India) for 48 h at 37°C, with shaking at 150 rpm. Bacteria were collected by centrifugation (4000xg, 15 min, 4°C) and resuspended in 20 mL of 10 mM Tris-HCl, pH 7.4. OMPs were extracted as described by Lopes et al. (2000) and Chansiripornchai (2004) and analyzed by Sodium Dodecyl Sulfate Poly Acrylamide Gel Electrophoresis (SDS-PAGE), using a 4% stacking gel and a 10% separating gel.
Purification of major OMPs from acrylamide gel: Extracted OMPs were electrophoresed in a 10% gel and visualized with 0.25 M KCl. Three major OMPs of 45, 53 and 70 kDa were cut from the gel and placed in separate dialysis tubes, containing 1.5 mL SDS-PAGE running buffer. The tubes were electrophoresed in a horizontal chamber for 30 min and dialyzed against PBS, overnight at 4°C. After dialysis, the gel pieces were discarded and tubes contents were concentrated with Nitrogen gas. The purified OMPs were analyzed by SDS-PAGE and quantified by dye-binding method.
Chickens: Day old commercial broiler chickens were procured from a breeder farm and grown until they became specific antibody negative (SAN). SAN chickens were used for production of antisera and also for preparing tracheal tissues for bacterial adherence inhibition assays. To ensure the chickens were free of maternal antibodies, they were tested by a commercial ORT-ELISA kit (IDDEX, USA) at days 1, 14 and 28.
Antisera: Antisera were prepared in SAN chickens. Total extracted OMPs or purified 45, 53 and 70 kDa OMPs were emulsified in ISA70 oil adjuvant at the ratio of 3 to 7 and injected subcutaneously to 4 weeks old SAN chickens (30 μg in a final volume of 0.5 mL/chicken). One more injection was given two weeks after the first immunization and the chickens were bled two weeks later. Sera were stored at -20°C until use. Antiserum against the whole cells of ORT was prepared as above, after inactivating the bacterial suspension with 3% of formalin at 60°C for 30 min.
Immunodot and Immunoblotting assays: In immunodot assay, antigen spots were made on small pieces of nitrocellulose membrane, using 10 μL volumes of the extracted total OMPs. The membranes were blocked in 5% skim milk in PBS, washed three times in PBS containing 0.05% Tween (PBS-T) and probed with 1/100 dilutions of sera. After washing as above, the membranes were incubated in a 1/200 dilution of a peroxidase conjugated goat anti-chicken IgG (KPL, USA) in PBS-T. The membranes were washed again and developed by 4-chloro-1-naphtol (Sigma, USA).
For immunoblotting, the extracted total OMPs were electrophoresed in a 10% gel and transferred to Nitrocellulose membrane. The membrane was cut in strips, blocked with skim milk and probed with the sera and conjugate as above.
Adherence studies: Adherence studies were performed as described by Gyimah and Panigraphy (1988), with some modifications. Two hundreds microliter volumes of the ORT suspension were incubated with 200 μL of each antiserum for 1 h at room temperature with occasional shaking. As a control, 200 μL of ORT suspension was incubated with 200 μL of a normal chicken serum. Each of the treated bacterial suspensions was added to 5 tracheal sections (1x3 mm rectangular sections), maintained in 4 mL of Krebs Ringer Tris saline (NaCl 7.5 g L-1, CaCl2 0.305 g L-1, MgSO4.7H2O 0.318 g L-1, KCl 0.383 g L-1, buffered with 0.05M Tris-HCl), pH 7.4, in a 50 mL conical tube. Bacteria and tissue sections were incubated for 1 h at 37°C in a shaker. After incubation, the tracheal sections were washed in three changes of PBS to dislodge non-adherent bacteria. Each tracheal section was ground in 2 mL of PBS using a sterile mortar and pestle and subjected to bacterial counting following two fold serial dilutions and plating on Columbia agar medium. Relative adherences were determined as follows and compared by students t-test
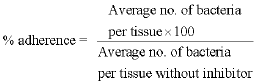 |
RESULTS
OMPs of an ORT serotype A were extracted and analyzed by SDS-PAGE and silver staining. The OMPs bands were in range of 32 to 83 kDa. Three major OMPs with apparent molecular weights of 45, 53 and 70 kDa were purified from the gel by electro elution (Fig. 1).
Antisera to ORT, total OMPs and each of the three eluted outer membrane proteins were prepared by subcutaneous injections of commercial broiler chickens. Two weeks after the second injection, the presence of relevant antibodies in the immune sera was demonstrated by immunodot but the control chickens and chickens immunized by 45 kDa OMP had not any ORT-reactive antibodies (data not shown).
 | |
| Fig. 1: | SDS-PAGE analysis of OMPs of ORT. Lane 1 to 6 represent the molecular weight markers, whole cell of ORT Serotype A, OMPs extracted from ORT, purified 70 kDa OMP, purified 45 kDa OMP and purified 53 kDa OMP respectively. The proteins were visualized by silver staining |
 | |
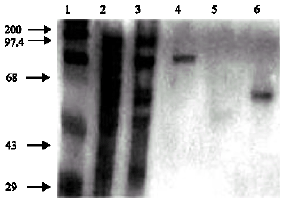
| Fig. 2: | Reactivity of antisera prepared against OMPs of ORT in immunoblotting. Lanes 1 to 4 represent the reactivity of sera against OMPs extracted from ORT, purified 70 kDa OMP, purified 45 kDa OMP and purified 53 kDa OMP respectively. Lane 5 indicates the reaction of normal chicken serum. Molecular weights are indicated at left |
The specificity of antigen-antibody reactions was confirmed by immunoblotting (Fig. 2). The immune sera against total OMPs, 53 and 70 kDa OMPs were capable of detecting respective antigens, but sera of control chickens and chickens immunized by 45 kDa OMP couldn't detect the OMPs antigens.
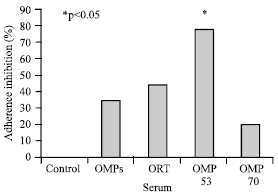 | |
| Fig. 3: | Inhibitory effects of antisera against OMPs of ORT on the adherence of ORT Serotype A to chicken tracheal epithelium. Inhibitory effects of normal chicken serum (control) and antisera against OMPs of ORT, whole cell of ORT and purified 53 and 70 kDa OMPs have been indicated from left to right, respectively |
The ability of immune sera and control serum to inhibit adherence of ORT to tracheal epithelial cells, has been shown in Fig 3. Serum against the 53 kDa OMP significantly (p<=0.05) showed the highest inhibition of adherence (relative adherence of 22 and 78% adherence inhibition). Sera prepared against the whole cell antigen of ORT, total OMPs and 70 kDa OMP inhibited the adherence of ORT by 44% (relative adherence of 56%), 34% (relative adherence of 66%) and 20% (relative adherence of 80%), respectively.
DISCUSSION
Adhesion of bacterial pathogens to epithelial cells is a key step in the establishment of most mucosal infections and thus an attractive target of infection intervention and prevention. Chansiripornchai (2004) has shown that lipopolysaccharide (LPS) of ORT inhibited bacterial adherence to human epithelial cell up to 90%, but the adherence of ORT to chicken epithelial tissues was not studied. On the other hand, OMPs of ORT may play a role in receptor (s) recognition, because the role of OMP (s) in adherence has been shown for many of bacteria like Heamophilus ducreyi (Dinitra et al., 2005), Actinobacillus pleuropneumoniae (Enriquez-Verdugo et al., 2004) and Haemophilus influenzae (Liu et al., 2004).
In the present work, we studied the probable role of ORT OMPs in adherence to chicken tracheal epithelium, based on the abilities of anti-OMPs antibodies to inhibit the bacterial adherence. The results of this study showed that antibodies raised against an OMP of 53 kDa could inhibit bacterial adherence up to 78%. Inhibitory effect of anti 53 kDa antibodies was significantly higher than those of antibodies produced against total OMPs and whole cell bacteria. The difference could be related to the amount of 53 kDa antigen in these preparations or the effect of other bacterial component (s) which might affect the immunogenicity of 53 kDa OMP. As we showed (Fig. 2 lane 1), the 53 kDa OMP was not detectable by antibodies prepared against the total OMPs.
Considering the results of this study, it can be concluded that OMPs of ORT could be implicated in bacterial adherence. Although Chansiripornchai (2004) hasn't used chicken trachea in his adherence studies, synchronous role of OMP (s) and LPS of ORT in attachment can be envisaged. In fact, ORT may recognize different receptors and bacterial OMP(s) and LPS may be implicated in the interaction with different kinds of receptors.
So far there have been a few studies on the OMPs of ORT. Lopes et al. (2000) investigated the use of OMPs of ORT in enzyme-linked immunosorbent assay (ELISA) to detect the exposure to ORT infection. They showed that OMPs-based ELISA was able to detect the exposure to ORT better than the agglutination test. Recently, in order to develop a cross protective subunit vaccine against ORT, Schuijffel et al. (2006) showed that immunization of chickens with a recombinant OMP of 32.9 kDa could reduce the respiratory pathology of ORT up to 74.6%. This OMP had been identified by screening of an ORT expression library with sera of live-vaccinated birds.
Based on the results we presented in this research, 53 kDa OMP could be a candidate for further studies in order to develop a recombinant vaccine against ORT.
REFERENCES
- Cauwerts, K., P. De Herdt, F. Haesebrouck, J. Vervloesem and R. Ducatelle, 2002. The effect of Ornithobacterium rhinotracheale vaccination of broiler breeder chickens on performance of their progeny. Avian Pathol., 31: 619-624.
Direct Link - Dinitra, C., I. Leduce, B. Oslen, C. Jeter, C. Harris and C. Elkins, 2005. . Haemophilus ducreyi outer membrane determinants, including Dsr A, define two clonal populations. Infection Immunity, 73: 2387-2399.
Direct Link - Enriquez-Verdugo, I., A.L. Guerrero, J.J. Serrano, D. Godinez, J.L. Rosales, V. Tenorio and M. Garza, 2004. Adherence of Actinobacillus pleuropneumoniae to swine lung collagen. Microbiology, 150: 2391-2400.
Direct Link - Gyimah, J.E. and B. Panigraphy, 1988. Adhesin-receptor interactions mediating the attachment of pathogenic Escherichia coli to chicken tracheal epithelium. Avian Dis., 32: 74-78.
Direct Link - Hafez, H.M., 2002. Diagnosis of Ornitobacterium rhinotracheale. Int. J. Poult. Sci., 1: 114-118.
Direct Link - Joubert, P., R. Higgins, A. Laperle, I. Mikaelian, D. Venne and A. Silim, 1999. Isolation of Ornitobacterium rhinotracheale from turkeys in Quebec, Canada. Avian Dis., 43: 622-626.
Direct Link - Liu, D., K.W. Mason, M. Mastri, M. Pazirandeh and D. Cutter et al., 2004. The C-terminal fragment of the internal 110-Kilodalton passenger domain of the Hap protein of nontypeable Haemophilus influenza is a potential vaccine candidate. Infect. Immunity, 72: 6961-6968.
Direct Link - Lopes, V., G. Rajashekara, A. Back, D.P. Shaw, D.V. Halvorson and K.V. Nagaraja, 2000. Outer membrane proteins for serologic detection of Ornitobacterium rhinotracheale infection in turkeys. Avian Dis., 44: 957-962.
Direct Link - Schuijffel, D.F., P.C.M. Van Emple, A.M.M.A. Pennings, Van J.P.M. Putten and P.J.M. Nuijten, 2005. Passive immunization of immune-suppressed animals: Chicken antibodies protect against Ornithobacterium rhinotracheale infection. Vaccine, 23: 3404-3411.
Direct Link - Schuijffel, D.F., R.P. Segers, P.C.M. Van Emple, J.P.M. Van Putten and P.J.M. Nuijten, 2006. Vaccine potential of recombinant Ornithobacterium rhinotracheale antigens. Vaccine, 24: 1858-1867.
Direct Link - Van Empel, P., H. Van den Bosch, P. Loeffen and P. Storm, 1997. Identification and serotyping of Ornithobacterium rhinotracheale. J. Clin. Microbiol., 35: 418-421.
Direct Link - Van Empel, P.C.M. and M.H. Hafez, 1999. Ornithobacterium rhinotracheale: A review. Avian Pathol., 28: 217-227.
Direct Link