
Nasreen Gill
Department of Zoology, University of Sindh, Jamshoro, Sindh, Pakistan
Azra A. Shaikh
Department of Zoology, University of Sindh, Jamshoro, Sindh, Pakistan
M. Munif Khan
Department of Zoology, University of Sindh, Jamshoro, Sindh, Pakistan
M. Saleh Memon
Institute of Biochemistry, University of Sindh, Jamshoro, Sindh, Pakistan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 24 | Page No.: 4479-4484







ABSTRACT
Present study was performed to determined the changes in the blood picture of the brown rats Rattus norvegicus naturally infected with intestinal cestodes. The results are based on the examination of 82 brown rats captured at random from Hyderabad city and its adjacent areas. Blood profiling of cestode (Hymenolepis diminuta and Hymenolepis nana) infected male and female rats showed significantly lower values of Total Erythrocyte Count (TEC), Haemoglobin (Hb) concentration, Haematocrit (Hct), Mean Corpuscular Haemoglobin (MCH) and Mean Corpuscular Haemoglobin Concentration (MCHC) as compared to their control (non-infected) rats of both sexes. The Mean Corpuscular Volume (MCV) value for infected male and female rats was found significantly higher than those of their control rats. A significantly higher Total Leukocyte Count (TLC) in male and female infected rats was noted as against the non-infected (control) rats of both sexes. The Differential Leukocyte Count (DLC) revealed significant increase in absolute number of each type of cell in cestode infected male and female rats. However, lymphocyte and basophil counts recorded from male infected rats were similar to the counts of their control rats. Conspicuous changes in the blood picture of cestode infected male and female rats indicate host-parasite interaction as well as the triggering of defense mechanism in the host against infection.
PDF Abstract XML References Citation
How to cite this article
Nasreen Gill, Azra A. Shaikh, M. Munif Khan and M. Saleh Memon, 2007. Influence of Intestinal Cestodes on the Blood Picture of the Brown Rats (Rattus norvegicus) of Hyderabad Sindh, Pakistan. Pakistan Journal of Biological Sciences, 10: 4479-4484.
DOI: 10.3923/pjbs.2007.4479.4484
URL: https://scialert.net/abstract/?doi=pjbs.2007.4479.4484
DOI: 10.3923/pjbs.2007.4479.4484
URL: https://scialert.net/abstract/?doi=pjbs.2007.4479.4484
INTRODUCTION
Maintenance of constant haematological values within certain limit is indispensable for each individual to perform normal physiological functions. Any alteration in such values may cause impaired metabolic activities in animals.
Cestodes, commonly known as tapeworms, inhabit the lumen of vertebrate’s intestine where they ate bathed in a veritable soup of nutrients which are absorbed through the tegument (Chappell, 1980; Cable, 1985). There are different ways in which parasites, including cestodes, inflict mechanical injury to their hosts (Chappell, 1980). Haematological as well as immunological responses to the helminthic infections have also been studied extensively in different hosts (Mathur and Johnson, 1989; Mohsin et al., 1991; Hunter et al., 2005; Torgerson, 2006; Raza et al., 2006; Sommerfelt et al., 2006; Smith et al., 2007).
Morphotaxonomy of endoparasites of rats has been the main subject of research in Pakistan (Bilqees and Siddiqui, 1981; Farooq and Yousuf, 1986; Faiyaz-ul-Haque et al., 1990; Fatima, 1991; Noor-un-Nisa, 2001). In past few studies and only in Pakistan, have been conducted to show the effects of different parasites (bladder worms and blood protozoans) on the haematological parameters of Rattus norvegicus (Gill et al., 2003; Gill and Khan, 2004, 2005). However, the influence of intestinal cestodes on the blood picture of Rattus norvegicus remains unstudied in Pakistan and elsewhere. Hence, the blood profiling of Rattus norvegicus naturally infected with intestinal cestodes seems vital and valid.
MATERIALS AND METHODS
In the present study (December, 2004-January, 2006) 82 rats (Rattus norvegicus), comprising 40 non-infected (86.36±2.78 g body weight) and 42 infected (85.76±2.38 g body weight) rats were captured at random from the Hyderabad city and its adjacent areas. They were kept (4 rats/cage) in the laboratory of Zoology Department, University of Sindh, Jamshoro. The rats were fed with food and water ad libitum.
In the text the term Infected refers to those rats, which harbour intestinal cestodes, where as non-infected means control rats.
Collection of cestodes: To avoid stressing effect on the body and other parameters, rats were dissected and examined within 24 h from the capturing time. Each animal was anaesthetized using chloroform or ether. Before dissection the sex and weight of each rat was noted. The intestinal cestodes were collected and stained by the methods describe by Schmidt (1988). Cestodes recovered from the intestine of Rattus norvegicus were thoroughly examined under the microscope and identified with the help of keys, related literature and description given by Yamaguti (1959) and Soulsby (1978).
Blood sampling and procedure: Blood was collected from the heart and immediately transferred into a sterilized vial containing a mixture of ammonium oxalate and potassium oxalate as anticoagulant (Wintrobe, 1961). The methods of Wintrobe (1961) were adopted for the determination of haematological values of Rattus norvegicus. Blood smears were prepared and stained with Giemsa’s stain for differential leukocyte counts (Schmidt, 1988).
Statistical analysis: All data were analyzed statistically using student’s t-test to determine significance difference between control and infected rats (Baily, 1981). Statistical significance was accepted at p≤0.05. Haematological values are expressed as Mean±SEM.
RESULTS
Identification: Cestodes recovered from the intestine of Brown rats Rattus norvegicus were identified as Hymenolepis diminuta and Hymenolepis nana.
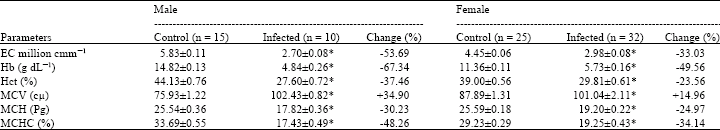
Erythrocytes indices during cestode infection: The erythrocyte indices (Table 1) were determined during cestode infection, caused together by Hymenolepis diminuta and Hymenolepis nana in the intestine of male and female rats (Rattus norvegicus). The mean values of TEC, Hb concentration, Hct, MCH and MCHC significantly declined (p<0.001) in infected male and female rats as compared to their controls rats of both sexes. Reduction in values of TEC, Hb concentration, Hct, MCH and MCHC was 53.69, 67.34, 37.46, 30.23 and 48.26%, respectively in male infected rats. While same erythrocyte indices were found to be decreased by 33.03, 49.56, 23.56, 24.97 and 34.14%, respectively in female infected rats. Mean Corpuscular Volume (MCV) estimation indicated significantly higher (p<0.001) values in male and female infected rats than their control rats. The elevation in MCV values was 34.90 and 14.96% in infected male and female rats, respectively.
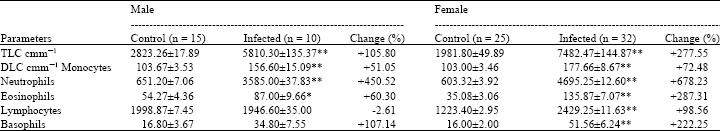
Leukocyte counts during cestode infection: The leukocyte counts during cestode infection in intestine of Rattus norvegicus are presented in Table 2. TLC were found significantly higher (p<0.001) during cestode infection in male and female rats by 105.80 and 277.55%, respectively than those of their control rats of both sexes. Amongst the differential leukocyte counts monocytes, neutrophils and eosinophils showed higher values at different levels of significance (Table 2) in infected male and female rats as compared to their control male and female rats. An increase of 51.05, 450.52 and 60.30% was recorded in monocyte, neutrophil and eosinophil counts, respectively in infected male rats,whereas counts increased by 72.48, 678.23 and 287.31% in the same type of cells, respectively in infected female rats.
| Table 1: | Erythrocyte indices (Mean±SEM), during of cestode infection in intestine of Rattus norvegicus |
 | |
| Significant: *: p<0.001, +: Increased, -: Decreased | |
| Table 2: | Leukocyte counts (Mean±SEM), during cestode infection in intestine of Rattus rvegicus |
 | |
| Significant: *: p < 0.01, **: p < 0.001, +: Increased, -: Decreased | |
As compared to the control female rats, significantly (p<0.001) increased values by 98.56 and 222.25% for lymphocyte and basophils, respectively were recorded from the infected female rats. However, male infected rats did not show any change in lymphocyte and basophil counts as compared to their control rats (Table 2).
DISCUSSION
Erythrocyte indices: During current observation the values of TEC, Hb concentration and Hct were found significantly (p<0.001) low by different percentage in male and female rats naturally infected with intestinal cestodes (Table 1). The reduction in TEC and Hb concentration was also noted by Reddy et al. (1989) in fish infected with haemoflagellate. Mathur and Johnson (1989) investigated blood profile of house rats (Rattus rattus) naturally infected with Vampirolepis fraternal (Cestoda) and observed lower TEC but significantly enhanced level of Hb. Significant reduction was noted in Hct (PCV) values in rats (Rattus rattus) naturally infected with Hymenolepis diminuta (Mathur and Johnson, 1987). Reduction in TEC, Hb concentration and Hct were also reported by Haroun et al. (1986), Chaudri et al. (1988) and Mohsin et al. (1991) due to Fasciola gigantica infection in different live stocks. Chaudhry et al. (1991) in Equines of Faisalabad (Pakistan) during Strongylus infection and Aliyu et al. (1997) in donkeys infected with Trypanosoma congolense reported significant fall in TEC, Hb concentrations and Hct values. Gill et al. (2003) and Gill and Khan (2005) also observed reduction in TEC, Hb concentration and Hct values during bladder worm (Cysticercus fasciolaris) and protozoan infections in Rattus norvegicus. Raza et al. (2006) noted decreased TEC and Hb concentration in sparrows infected with Heterakis gallinea and Ascaridia gallinae. Finding of present study as well as above observations clearly indicate reduction in the values of TEC, Hb concentration and Hct due to parasitic infections in different types of hosts.
The percentage change (drop) in TEC as compared with the control rats of both sexes provides sufficient evidence for the abnormal results in term of low counts of erythrocytes (Table 1). The fall in TEC recorded from the infected male and female rats may be attributed to the blood loss (various types of anaemia and haemorrhage), erythropoietin deficiency, hemolysis (erythrocyte destruction), malnutrition (nutritional deficiency of iron, folic acid, vitamin B12 or vitamin B6) or some other factors (Milone, 2004; Nordenson, 2004). Since the erythrocytes are rich in haemoglobin, hence the reduction in TEC recorded during current study may be one of the reasons for the decrease in Hb concentration during cestode infection in rats of both sexes. Decreased Hb concentration in the infected rats, as indicated by % change in Table 1, may also be associated with the blood loss and various types of anaemia (Milone, 2004). The decreased Hct values, obtained from the infected male and female rats, may be due to anaemia (Anonymous, 1979; Milone, 2004) which is also supported by the fact that the values of TEC and Hb concentration were found low in the infected rats of both sexes (Table 1).
This study showed significantly enhanced value of MCV by 34.9 and 14.96% in male and female infected rats, respectively. Haroun et al. (1986) and Mohsin et al. (1991) also observed increased values of MCV due to fascioliasis in ovine and cattle, respectively however no significant differences were observed between the young adult infected and control animals. Swarup et al. (1986) and Chaudri et al. (1988) also did not find any significant difference between normal and Fasciola infected animals. It has been indicated by Mohsin et al. (1991) that the MCV values usually rise in deficiency of Vitamin B12, folic acids, niacin (Pellagra preventing factor) and cobalt. The deficiencies of these nutrients, as suggested by Mohsin et al. (1991), may be due to parasitic infection (Fascioliasis). In this finding the observed elevation in the MCV value of cestode infected male and female rats can be attributed to the macrocytic anemia resulting by the deficiency of B12, B6, folic acids, niacin and cobalt (Mohsin et al., 1991; Cutler, 2002; Nordenson, 2004).
Cestode infection revealed significant reduction in MCH values by 30.23 and 24.97% in male and female rats, respectively. This infection also showed similar effect on MCHC values by causing significant fall by 48.26 and 34.14% in male and female rats, respectively. It has been suggested that due to the use of RBC in calculation, MCH is not as accurate as MCHC in the diagnosis of severe anaemia (Anonymous, 2004). Low MCH and MCHC indices, recorded during current study, indicate hypochromic anaemia in cestode infected rats. Blood loss as well as vitamin B6 and iron deficiency can be accounted for the hypochromic anaemia during cestode infection in rats of both sexes (Cutler, 2002; Anonymous, 2004; Nordenson, 2004).
Total leukocyte count: In the present study, TLC was found significantly high during cestode infection in both sexes of rats indicating 105.80 and 277.55% rise as compared to the control. Mathur and Johnson (1989), while working on the blood profile of Rattus rattus naturally infected with Vampirolepis fraterna (Cestoda), observed significantly enhanced level of leukocytes. Significant increase in TLC has also been reported during Fasciola infection in cattle (Mohsin et al., 1991), haemoflagellate infection in fish (Reddy et al., 1989), Trypanosoma congolense infection in donkeys (Aliyu et al., 1997), bladder worm (cestode) infection in liver of Rattus norvegicus (Gill et al., 2003), protozoan infection in blood of Rattus norvegicus (Gill and Khan, 2004) and Heterakis gallinea and Ascaridia gallinae infection in sparrows (Raza et al., 2006). In contrast to these reports Chaudhry et al. (1991) observed slight reduction in TLC but it remained with in the normal range during Strongylus infection in Equines. Leukocytes are responsible for the defense of the organism and protection of the body against foreign invaders including parasites (Macer, 2003). Leukocytosis at higher percentage (105.80 and 277.55%) in infected rats of both sexes (Table 2) therefore suggests immune response of the host against infection.
Differential leukocyte count: Differential leukocyte count was performed to examine the variations in absolute number of each type of leukocytes during cestode infection in the intestine of Rattus norvegicus. Each type of leukocytes has its own normal values for every individual. The functions of the different types of leukocyte vary, but generally their most important functions are the defense of the body against infection and to produce or at least transport and distribute antibodies in the immune response (de Gruchy, 1976; Anonymous, 2004).
Increased monocyte, neutrophil and eosinophil counts have been reported in animals harbouring different types of parasites (Reddy et al., 1989; Mohsin et al., 1991; Gill and Khan, 2004; Sommerfelt et al., 2006). Decreased eosinophil and lymphocytes was also reported during Heterakis gallinea and Ascaridia gallinae infection in sparrows (Raza et al., 2006).
Monocytosis commonly occurs during inflammation, bacterial and different types of infections (de Gruchy, 1976; Macer, 2003; Inoue, 2004). This finding also showed monocytosis in cestode infected male and female rats, Rattus norvegicus. This observation (monocytosis) may be associated with the infection as well as the defense mechanism of the host.
Neutrophils play an important role in the defense and protection of the body against diseases and infections (de Gruchy, 1976; Daniels, 2002). Daniels (2002) pointed out a number of possible causes, including haemolytic anaemia for an abnormally high neutrophil counts. In this study differential leukocyte counts revealed remarkable higher values of neutrophil counts by 450.52 and 678.23% in male and female infected rats, respectively than those of their control rats of both sexes. The haemolytic anemia, as indicated by Daniels (2002), may be one of the causes for such an abnormally high neutrophil counts in male and female infected rats. Though the type of anemia noted in this study is not evident, however the anaemic condition is supported by the fact that both total erythrocyte count and haemoglobin concentration were low in the infected rats of both sexes (Table 1). Another possible reason that seems to increase neutrophils in the blood of infected male and female rats over their normal (control) rats is the triggering of the immunological phenomenon or the enhancement in the potentialities of the defense mechanism in the host (de Gruchy, 1976; Daniels, 2002).
It has been indicated that the eosinophils are used by the body to protect against allergic reactions and parasites (Anonymous, 2004). Teo et al. (1985), Nutman et al. (1987) and Weller (1992) also demonstrated that parasitic disease is the principal cause of eosinophilia in many nonindustrialized parts of the world. In established parasitic infections only moderately high blood eosinophil counts are found, but in the early stages of infection when parasites are migrating through tissues very high counts were noted (Weller, 1992; Auletta and Shurin, 2000). The infectious organisms most likely to cause eosinophilia are parasites (helminthes, protozoan), fungi, few viruses and bacteria (de Gruchy, 1976; Auletta and Shurin, 2000; Abramson and Melton, 2000). The eosinophilic response noted during cestode infection in rats of both sexes may be attributed to the tissue migration cycle of the parasites (Weller, 1992; Auletta and Shurin, 2000) as well as the body protection against allergic reactions and parasites (Anonymous, 2004).
Lymphocytes are the main constituents of the immune system, which is a defense against the attack of pathogenic micro-organisms. Reddy et al. (1989) recorded significant elevation in small lymphocyte counts during haemoflagellate infection in fish. Gill et al. (2004) reported lymphopenia and lymphocytosis during bladder worm infection in liver of male and female rats (Rattus norvegicus) respectively. Whereas, protozoan infection in blood of Rattus norvegicus of both sexes caused only lymphocytosis (Gill and Khan, 2004). In contrast to these observations, Mohsin et al. (1991) did not show any significant change in lymphocyte count in cattle harbouring Fasciola gigantica. Present study demonstrated lymphocytosis only in female infected rats which can be attributed to the development of defense power in the body. However no change could be observed in lymphocyte count during cestode infection in male rats. It seems that such infection did not trigger the immunological phenomena in male rats.
Basophils are hypersensitive effector cells and perform important role in host defense mechanisms, in innate immunity against pathogenic organisms and allergic reactions (Falcone et al., 2000). The level of basophils can be high in response to a number of causes and factors such as viral infection, haemolytic anaemia, toxic anaemias of long standing, myxoedema, hypothyroidism, ovulation, some conditions that cause inflammation and increased estrogen (de Gruchy, 1976; Abramson and Melton, 2000; Daniels, 2002). Cestode infection did not show any change in basophil count in male rats as compared to their control. This observation is in agreement to those of Mitre and Nutman (2003) who reported no relationship between parasitic infection and peripheral basophilia in humans. In contrast to male infected rats, this study demonstrated basophilia in females during cestode infection. Reddy et al. (1989) also noticed slight increase in the number of basophils in the fishes infected with haemoflagellate Trypanosoma vittai. However, these workers did not mention the sex of the fishes. A number of causes and factors have been shown for basophilia (de Gruchy, 1976; Abramson and Melton, 2000; Daniels, 2002) but at present it is not possible to assign a definite cause or factor which results in the elevation of basophils in the infected female rats. However, increased estrogen and ovulation may be the possible reasons for the basophilia in infected female rats (Abramson and Melton, 2000; Daniels, 2002).
It is now concluded that there exists a relationship between haematological indices and cestode infection, which is supported by the changes, detected in the blood picture of Rattus norvegicus. The information derived from this study is extremely valuable to elucidate host-parasite interaction as well as immune response against the invasion and establishment of the parasite in the host. This study is conducted for the first time from Rattus norvegicus captured from the study areas.
REFERENCES
- Abramson, N. and B. Melton, 2000. Leukocytosis: Basic of clinical assessment. Am. Fam. Physician, 62: 2053-2060.
Direct Link - Auletta, J.J. and S.B. Shurin, 2000. Eosinophilia: What does it mean? Contemp. Pediatr., 17: 61-61.
Direct Link - Falcone, F.H., H. Haas and B.F. Gibbs, 2000. The human basophil: A new appreciation of its role in immune responses. Blood, 96: 4028-4038.
CrossRefDirect Link - Gill, N. and M.M. Khan, 2004. Effects of blood protozoans on the haematological parameters of Rattus norvegicus of Hyderabad Sindh, Pakistan. Proc. Parasitol., 37: 17-25.
Direct Link - Haroun, E.M., H.A. Gadir and A.A. Gomel, 1986. Studies on naturally occurring ovine fascioliasis in Sudan. J. Helminthol., 60: 47-53.
PubMedDirect Link - Hunter, M.M., A. Wang, C.L. Hirota and D.M. McKay, 2005. Neutralizing anti IL-10 antibody blocks the protective effect of tape worm infection in Murine model of chemically induced colitis. J. Immunol., 174: 7368-7375.
Direct Link - Mitre, E. and T.B. Nutman, 2003. Lack of basophilia in human parasitic infections. Am. J. Trop. Med. Hyg., 69: 87-91.
Direct Link - Nutman, T.B., E.A. Ottesen, S. Ieng, J. Samuels and E. Kimball et al., 1987. Eosinophilia in Southeast Asian refugees: Evaluation at a referral center. J. Infect. Dis., 155: 309-313.
PubMedDirect Link - Raza, S.A., M. Athar Khan, M. Younus, I. Khan and T. Abbas, 2006. Prevalence of helminthiasis, its hematology and treatment with Fenbendazole in the migratory sparrows (Alauda arvensis). Proceedings of the 3rd National Conference on Agriculture, Animal Science, November 21-23, 2006, Sindh Agriculture Univ. Tandojam, pp: 41.
- Smith, P., N.E. Mangan, C.M. Walsh, R.E. Fallon, A.N.J. McKenzie, N.V. Rooijen and P.G. Fallon, 2007. Infection with helminthic parasite prevents experimental colitis via a macrophage-mediated mechanism. J. Immunol., 178: 4557-4566.
Direct Link - Sommerfelt, I.E., G. Santillan, G. Mira, M. Ribicich, A. Betti and R. De-Torres, 2006. Toxocara canis infection in a pig model: Immunological, Haematological and Blood biochemistry. J. Helminthol., 80: 73-77.
Direct Link - Teo, C.G., M. Singh, W.C. Ting, Y.W. Ong and L.C. Seet, 1985. Evaluation of the common conditions associated with eosinophilia. J. Clin. Pathol., 38: 305-308.
PubMedDirect Link - Torgerson, P.R., 2006. Canid immunity to Echinococcus sp. Impact on transmission. Parasite Immunol., 28: 295-303.
Direct Link