
Zahedi- Zahra
University of Tarbiyat Modares, Tebran, Iran
S. Esmaeelkhanian
Department of Biotechnology, Animal Science Research Institution of Iran, Tehran, Iran
R. Vaez Torshizi
University of Tarbiyat Modares, Tebran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 24 | Page No.: 4455-4460







ABSTRACT
Polymorphic loci, genetic parameters (Polymorphic Information Content, heterozygosity, Shannon Index, the number of actual alleles, the number of effective alleles) and genetic variation within Balouchi sheep population located in Abbasabad station of Mashad, was assessed using 12 microsatellite markers. The Polymerase Chain Reaction (PCR) reactions were successfully done with all primers except for OarHH62 and OarAE64, one locus (CSSM059) was monomorph, whereas the other 9 loci containing CSSM018, CSSM006, BMS1004, BMS2361, BMS744, BMS799, BMS1316, BMS2213 and TGLA122 were polymorph with 6, 6, 4, 4, 5, 4, 2, 3 and 3 number of alleles, respectively. In this population, heterozygosity varied from 0.5 to 0.72; among 9 polymorphic loci, BMS1316 and BMS2361 had the most and the least diversity, respectively. The study of Shannon Index and Polymorphic Information Content (PIC) indicate the least and the most values for BMS1316 and BMS2213 loci, respectively. The most number of actual alleles were belong to CSSM018 and CSSM006 loci with 6 alleles and the least was belong to BMS1316 locus with 2 alleles. BMS2361 locus had the most and BMS1316 locus had the least number of effective alleles. All loci were at Hardy-Weinberg equilibrium (p< 0.005). In this population, average heterozygosity was estimated as 0.54. In general, it can be concluded that Balouchi sheep population of Abbasabad station in Mashad (in Iran) has approximately high genetic diversity with respect to the studied microsatellites and microsatellite markers have approximately high polymorphism and therefore can be used for genetic studies.
PDF Abstract XML References Citation
How to cite this article
Zahedi- Zahra, S. Esmaeelkhanian and R. Vaez Torshizi, 2007. Microsatellite Variation in One Breed of Iranian Sheep with 12 Markers. Pakistan Journal of Biological Sciences, 10: 4455-4460.
DOI: 10.3923/pjbs.2007.4455.4460
URL: https://scialert.net/abstract/?doi=pjbs.2007.4455.4460
DOI: 10.3923/pjbs.2007.4455.4460
URL: https://scialert.net/abstract/?doi=pjbs.2007.4455.4460
INTRODUCTION
Genetic variation is essential for the genetic improvement of domestic animals. In fact, variation is the raw material on which the breeder works, so we should conserve our genetic resources. The realization of the need for conservation of animal genetic resources is not new and in fact has been on the international agenda for some 50 years (Barker, 1994). Translation of this realization into action has been slow, although there have been notable achievements and recent developments are encouraging. The development of molecular biological techniques has created new possibilities for the selection and genetic improvement of livestock, the discovery of the Polymerase Chain Reaction (PCR) had a major impact on the research of eukaryotic genomes and contributed to the development and application of various DNA markers. DNA markers like microsatellites have a potential application over a relatively broad field in animal breeding and genetics. The technique has direct practical application for livestock breeders. Microsatellites are used widely in construction of linkage maps in many species. High levels of polymorphism couped with the ease of analysis of the PCR have resulted in this type of marker being one of the most widely used for genetic analysis. A large number of highly polymorphic microsatellites, including sheep, cattle and other ruminants (Gortari et al., 1997; Hayes et al., 1996; Jenkins et al., 1997) facilitating the use of these markers in researches. Preservation of genetic diversity is important for conservation of biological diversity and utilization of genetic resources for economic and environmental sustainability. The focus of conservation efforts in animal breeding should be expanded to encompass all biodiversities. Conservationists also use various genetic markers in evolutionary and genetic biodiversity studies. In conclusion, the total biodiversity on earth represents a resource that should be conserved to supply genetic diversity for animal improvement. Breed conservation is required to cope with changes in consumer preference or productive environment, to supply genes for disease resistance and other simply inherited characteristics (Masone, 1996). In Iran, sheep is a main domestic animal with population about 60 millions. So, evaluation of genetic variation in Iranian sheep is a critical necessity. The objective of this study is to describe the native sheep breed of Iran to assess polymorphism within the Balouchi sheep population of Abbasabad station of Mashad using 12 microsatellite markers and to investigate of the studied microsatellite markers offer enough variability for further studied especially for Marker Assisted Selection (MAS). Stone et al. (1995) presented primer sequences and genotyping information for 390 informative bovine microsatellite markers; Gortari et al. (1997) reported the evaluation of 1036 bovine microsatellite primer pairs for their suitability as linkage markers in sheep and Maddox et al. (2001) proposed an enhance linkage map of the sheep genome comprising more than 1000 loci.
MATERIALS AND METHODS
Animals: Like the dog, sheep is one of the first animals that has been domesticated in Iran. The history of sheep and specially all the livestock in Iran, back to thousands of years ago. The first fleece of sheep is dated back to 5000 BC in the form of a crude clay image of a sheep found in Sarab of Iran. It is almost the most rife livestock in this country too. In 1986 the Iranian sheep population was estimated at 34500000 head. The Balouchi is a breed of sheep that originated in the eastern (Khorasan province) and south eastern area (Sistan-o-Balouchestan) and Kavir Markazi of Iran, Balouchi sheep is a widespread breed of meat-wool sheep, well adapted to a wide range of harsh environmental conditions in eastern Iran, one of the arid subtropical areas of the world. They are fat-tail sheep using for dairy, meat and wool (with good quality carpet wool), they are white with black marks on the head and legs, on the other hand, the fleece is white with pigmented head and legs. The wool is coarse with modulation. Body size varies between 35 and 40 kg in adult ewes, milk yield between 40 and 50 kg in a period of about 125 days. Rams can be either horned or polled and ewes are polled. This breed accounts for 50% of the national flock in Iran. Some other traits and characteristics are shown in Table 1. This study has been done in Iran in spring 2006.
Samples: Blood samples were obtained from 160 individuals and every 160 individuals were genotyped for 12 microsatellite markers. Blood samples of 10 mL were collected in EDTA tubes and frozen at -20°C. DNA was isolated using the salting-out method (Miller et al., 1988). Approximately, 150 ng DNA was used as template for PCR. All 160 sheep samples were genotyped with 12 microsatellite markers: CSSM018, CSSM059 (Maddox et al., 2001; Moore et al., 1994), CSSM006 (Looft et al., 1994; Moxon and Wills, 1999), BMS1004, BMS2361, BMS744, BMS792, BMS2213 (Maddox et al., 2001; Stone et al., 1995), BMS1316 (Gortari et al., 1997; Maddox et al., 2001; Stone et al., 1995), TGLA122 (Georges and Massey, 1992; Gortari et al., 1997; Maddox et al., 2001), OarHH62 and OarAE64 (Davis et al., 1996; Ede et al., 1994; Maddox et al., 2001).
The microsatellite loci were amplified by PCR for all animals. All primer sets were amplified in separated reactions. Amplification was carried out in a total volume of 15 μL containing the following constitutes: 150 ng of DNA used as a template, PCR buffer 1 X, 200 μm of each dNTP, 0.25 μm of each primer, 1 unit of Taq polymerase and 4 μm of MgCl2. PCR amplifications were performed by an initial denaturation of 4 min at 95°C, followed by 35 cycles of 30 sec of denaturation at 95°C, 30 sec annealing at different temperatures and 45 sec of extention at 72°C and the final extention step was lenghted 5 min at 72°C to ensure that polymerization was complete. The PCR were performed using Eppendorf and Biometra (T-gradiant) thermocyclers. The amplified products were run and electrophoresed on 8% denaturing polyacrylamide gels to confirm amplification and the DNA bands were visualized by Rapid Silver Staining (Bassam and Cateano-Anolles, 1993). Genotype of individuals estimated from the gels, gene and genotype frequency obtained by direct counting. Once genotypes were determined, allelic frequencies were calculated. Allelic frequencies for each primer in this population were computed simply by dividing the counts for each allele by the total number of alleles found in that population (for that certain primer). Chi-square analysis was employed to evaluate differences in allelic frequencies. Hardy-Weinberg equilibrium was evaluated by Chi-square (χ2T) and G2T test. Average heterozygosity (gene variability) per locus was calculated using the formula:
| r | = | The number of loci assessed, |
| Hk | = | Heterozygosity content for locus k (Hedrick, 1999). |
| Table 1: | Some productive traits and visual characterizations of Balouchi sheep |
 | |
For better assessed of genetic variation we used Shannon's Information Index (I) by the formula:
Polymorphic Information Content (PIC) per locus was calculated using the formula:
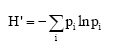 |
| k | = | The number of alleles, |
| pi and pj | = | Show frequency, |
| i and j | = | Allele, on this population, respectively (Buchanan and Thun, 1998). |
Hardi-Weinberg equilibrium, observed and expected heterozygosity, Shannon index and polymorphic parameters (the number of actual alleles and the number of effective alleles) were calculated using POPGENE computer program (Yeh et al., 1999) and PIC was calculated using PIC computer program (Ott, 1992).
RESULTS AND DISCUSSION
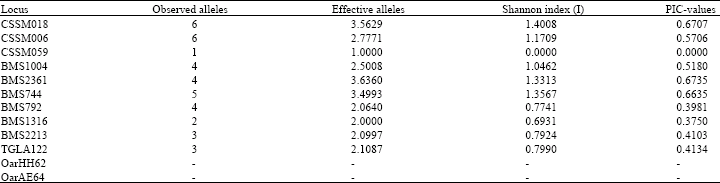
In this research, 12 microsatellites and the PCR reactions were optimized for all 12 primers. Allele size, allele range, the number of observed and effective alleles, Shannon Index (I), PIC value, observed heterozygosity (Hobs) and expected heterozygosity (Hexp) were calculated. Amplifications were successfully done for all primers except for OarHH62 and OarAE64. CSSM059 locus was monomorph for this breed (Balouchi), whereas the other 9 loci, containing CSSM018, CSSM006, BMS1004, BMS2361, BMS744, BMS792, BMS1316, BMS2213 and TGLA122 were polymorph with different number of observed alleles that are presented in Table 4. Generally, a total of 38 alleles were detected in the 12 studied loci. The mean number of alleles, calculated from 10 microsatellite, 9 microsatellite loci were polymorph with the number of alleles per locus ranging from 2 (BMS1316) to 6 (CSSM018, CSSM006). The following are results obtained for each locus, briefly:
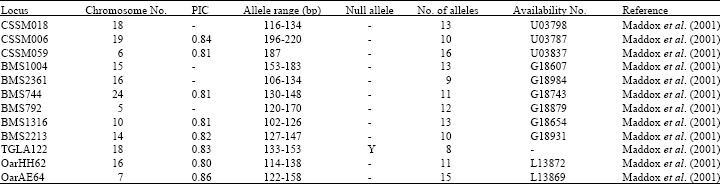
CSSM018 and CSSM006 loci showed the most number of alleles (6 alleles) (Table 3), for locus CSSM018 the allele range in this study was between 122 to 152 base pairs, whereas in previous research was 116-134 bp, so CSSM018*136, CSSM018*142 and CSSM018*152 alleles were the new one in this research and CSSM006*228, CSSM006*232 and CSSM006*240 alleles were reported for the first time. Allele range for locus CSSM006 (200-240 bp) was according to allele range obtained with Maddox et al. (2001) 196-220 bp. Generally, results obtained for loci CSSM018 and CSSM006 were according to Maddox et al. (2001) and Gortari et al. (1997) and CSSM018 according to Maddox et al. (2001) Moore et al. (1994) and for CSSM006 studied by Maddox et al. (2001), Looft et al. (1994) and Moxon and Wills (1999). According to Maddox et al. (2001) CSSM059 showed 16 alleles, but in our research, this locus was monomorph with allele size 211 bp and so with zero PIC value and Shannon Index (Table 2, 3, 5). CSSM059*211 has not been reported in other studies yet (Maddox et al., 2001; Moore et al., 1994). BMS1004 new alleles for this locus was BMS1004*187 (Table 2). This locus showed 4 alleles in this population, with PIC-value about 0.52 (Table 3), allele range obtained in this study was near to the previous studies (Maddox et al., 2001; Stone et al., 1995). Locus BMS2361 showed the highest PIC-value and as a result the highest heterozygosity, also has high Shannon Index after the locus CSSM018 (Table 3, 4). Two new alleles found in this locus was BMS2361*138 and BMS2361*154 (Table 2). BMS2361 showed 4 alleles in this population and has the highest effective alleles among 10 loci (Table 3) (Maddox et al., 2001; Stone et al., 1995) for locus BMS744 the number of allele and the allele range obtained for this locus is not completely according to earlier studies, it's probably because the population and the breed is different from the previous studies; in this locus three new alleles BMS744*154, BMS744*166 and BMS744*170 were found (Table 2, 5) (Maddox et al., 2001; Stone et al., 1995). Allele range for locus BMS792 was between 134-154 bp and was according to the previous researches (Maddox et al., 2001) There was any new allele in this locus (Table 2, 5) (Maddox et al., 2001; Stone et al., 1995) for locus BMS1316, 2 alleles with 2 effective alleles are reported in this research and have been reported in other studies. Maddox et al. (2001) reported 13 alleles for locus BMS1316 (Table 2, 5), in this population this locus has the lowest values of PIC, Shannon Index and heteroxygosity (Table 3, 4) (Gortari et al., 1997; Maddox et al., 2001; Stone et al., 1995) for locus BMS2213, the alleles BMS2213*153 and BMS2213*167 have not been reported before (Maddox et al., 2001) reported 10 alleles for this locus and in this population it shows just 3 alleles (Table 2, 5) (Maddox et al., 2001; Stone et al., 1995). TGLA122*171 and TGLA122*217 reported here for the first time (Table 2).
| Table 2: | List of the primers used with allele sizes |
 | |
| *: New alleles have been found in this population | |
| Table 3: | The number of actual and effective alleles, PIC-values and Shannon Index (I) per locus |
 | |
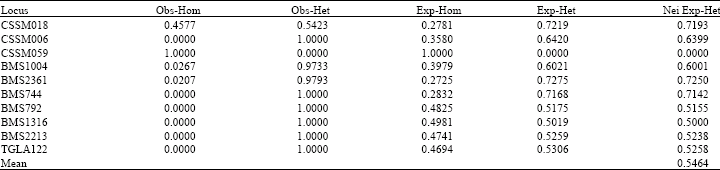
| Table 4: | Observed and expected heterozygosities, obs., exp. Homozygosities and Nei expected heterozygosities are presented |
 | |
The number and the range of alleles for locus TGLA122 were in contrast with previous studies, the highest null allele were detected for this locus (Georges and Massey, 1992; Gortari et al., 1997; Maddox et al., 2001).
In the summary of above explanation, results obtained for loci CSSM018 and CSSM006 were according to Maddox et al. (2001) and Gortari et al. (1997). Allele range for locus CSSM006 (200-240 bp) was according to allele range obtained with Maddox et al. (2001) 196-220 bp. The number and the range of alleles for loci BMS744 and TGLA122 were in contrast with the previous studies (Stone et al., 1995) (Table 2, 3). Comparing the number, the size and the range of alleles obtained in this study, new alleles, especially on loci CSSM006, BMS792, BMS2213 and TGLA122, were found at Iranian sheep that have not been previously reported and some alleles that have reported in other studies, are not obtained in this study (Moore et al., 1994). CSSM006*228, CSSM006*232, CSSM006*240 alleles, BMS1004*187 allele, BMS2361*138, BMS2361*154 alleles, BMS744*154, BMS744*166, BMS744*170 alleles, BMS2213*153, BMS2213*167 alleles, TGLA122*171 and TGLA122*217 alleles are being described here for the first time. Neither of these alleles have been previously reported in the literature; this shows that Iranian sheep has several new alleles on 8 polymorph loci. Seventeen previously unreported alleles were found during the course of this experiment. CSSM006-220, BMS2361-106 and BMS1316-126 alleles are being described in other studies and not in this experiment (Table 2). According to the previous studies, null allele has just been reported for locus TGLA122, but here, we observed null allele for all loci especially for locus TGLA122, it's probably shows that mutation has been accured in the place of the primers on the genome of Balouchi sheep and the mutation has been fixed in this population. Allele frequencies estimated from genotypes for each locus are shown in Table 2. Low variability was found in BMS1316, with only 2 alleles, but CSSM018 and CSSM006 with 6 alleles presented high levels of diversity. In conclusion, this work has revealed that genetic diversity exist among the Balouchi sheep population studied. Genetic variability parameters are presented in Table 3 and 4. Mean heterozygosity (0.54) showed high diversity levels in this sheep.
| Table 5: | Loci characterization reported in previous studies |
 | |
Heterozygosity and PIC estimates are normally correlated and as expected based on gene variability, BMS2361 showed the highest and BMS1316 the lowest values for both parameters (Table 3, 4). The highest and the lowest values of PIC for this breed (Balouchi) ranged from 0.3750 (BMS1316) to 0.6735 (BMS2361). Maddox et al. (2001) reported PIC = 0.81 for locus CSSM059, whereas in this study, this locus was monomorph with PIC = 0.00; according to Maddox et al. (2001) PIC values for loci CSSM006, BMS744, BMS1316, BMS2213 and TGLA122 were 0.84, 0.81, 0.81, 0.82 and 0.83, respectively and in this experiment we obtained the values 0.57, 0.66, 0.37, 0.41 and 0.41, respectively for mentioned loci. Comparing the PIC-values of primers in this study with the previous ones show large differences between PIC-values and also between the numbers of alleles; as mentioned, it's because of the different population that studied. Each primer has certain value of PIC in each population or in each breed, so we should obtain the PIC-value for each primer in certain population and the PIC-values of primers in previous studies can lead us to choose the primers that are more polymorph. For all 9 polymorphic loci, the mean expected heterozygosity was 0.54, while for individual locus average Hexp varied from 0.5019 (BMS1316) to 0.7275 (BMS2361) (Table 4). All loci were at Hardy-Weinberg equilibrium (p<0.005), it's probably because of the random mating that has been done in this population. In general, it can be calculated that Balouchi sheep population of Abbasabad station in Mashad has approximately high genetic diversity with respect to studied microsatellites and microsatellite markers have approximately high polymorphism and therefore can be used for genetic studies; on the other hand, microsatellite is an useful tool for evaluating of variation in this population.
REFERENCES
- Bassam, B.J. and G. Cateano-Anolles, 1993. Silver staining of DNA in polyacrylamide gels. Applied Biochem. Biotechnol., 42: 181-188.
Direct Link - Davis, K.P., J.F. Maddox, D.I. Hulme and K.U. Beh, 1996. Ovine dinucleotide repeat polymorphism at four anonymous loci. Anim. Genet., 27: 382-383.
PubMedDirect Link - Ede, A.J., C.A. Pierson, H.M. Henry and A.M. Crawford, 1994. Ovine microsatellites at the OarAE64, OarHH22, OarHH56, OarHH62 and OarVH4. Anim. Genet., 25: 51-52.
Direct Link - De Gortari, M.J., B.A. Freking, S.M. Kappes, K.A. Leymaster, R.T. Stone, C.W. Beattie and A.M. Crawford, 1997. Extensive genomic conservation of cattle microsatellite heterozygosity in sheep. Anim. Genet., 28: 274-290.
CrossRefDirect Link - Hayes, H., C. Le Chalony, G. Goubin, D. Mercier, E. Payen, C. Bignon and K. Kohno, 1996. Localization of ZNF164, ZNF146, GGTA1, SOX2, PRLR and EEF2 on homologous cattle, sheep and goat chromosomes by fluorescent in situ hybridization and comparison with t he human gene map. Cytogenet. Cell Genet., 72: 342-346.
Direct Link - Jenkins, Z.A., H.M. Henry, S.M. Galloway, K.G. Dodds and G.W. Montgomery, 1997. Comparative linkage mapping of genes on sheep chromosome 3 provides evidence of chromosomal rearrangements in the evolution of bovidae. Cytogenet. Cell Genet., 78: 272-274.
Direct Link - Maddox, J.F., K.P. Davis, A.M. Crawford, D.J. Hulme and D. Viaman et al., 2001. An enhance linkage map of the sheep genome comprising more than 1000 loci. Genome Res., 11: 1257-1289.
Direct Link - Mason, I.L., 1996. A World Dictionary of Livestock Breeds, Types and Varieties. 4th Edn., CAB International, Wallingford, UK., Pages: 273.
Direct Link - Miller, S.A., D.D. Dykes and H.F. Polesky, 1988. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res., 16: 1215-1215.
PubMedDirect Link - Moore, S.S., K. Byrne, K.T. Berger, W. Barendse, F. McCarthy, J.E. Womack and D.J. Hetzel, 1994. Characterization of 65 bovine microsatellites. Mamm. Genome, 5: 84-90.
Direct Link - Moxon, E.R. and C. Wills, 1999. DNA microsatellites: Against of evolution. Sci. Am., 271: 94-99.
PubMedDirect Link - Stone, R.T., J.C. Polido, G.M. Duyk, S.M. Kappes, J.W. Keele and C.W. Beatti, 1995. A small-insert bovine genomic library highly enriched for microsatellite repeat sequences. Mamm. Genome, 6: 714-724.
CrossRefDirect Link