
Leila Roushangar
Center of Drug Applied Research, Tabriz University of Medical Sciences, Iran
Jafar Soleimani Rad
Department of Histology and Embryology, Tabriz University of Medical Sciences, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 24 | Page No.: 4413-4419







ABSTRACT
Humans are continuously exposed to harmful environmental factors, including Electromagnetic Fields (EMF) produced by home appliances, diagnostic tools and industrial instruments. The present study investigates morphologically the effects of EMF on ovarian follicles. Female Wistar rats were exposed to 3 mili Tesla EMF for 4 h day-1 for 4 months, the ovaries from both experimental and control groups were processed for Transmission Electron Microscopy (TEM) and TUNEL reaction technique. TEM revealed that in the oocytes from experimental group the nuclei were shrunken and zona pellucida appeared narrower than those in the control group. The number of microvilli were decreased significantly and in their cytoplasm there were several lipid droplets and organels were dispersed. Signs of apoptosis such as condensed nuclei, chromatin margination and nuclear membrane dilatation were common in granulosa and corona radiata cells from the EMF-exposed rats than those from the control group. Primary morphological changes in the granulosa cells from this group were retraction of granulosa cells, loss of microvilli and condensation or loss of mitochondrial cristae. The morphological changes in oocytes seem to indicate the cytotoxic effect of EMF and the changes occurred in granulosa cells coincide with initiation of apoptosis in granulosa cells. Present results suggest that EMF exposure may interfere with normal process of folliculogenesis.
PDF Abstract XML References Citation
How to cite this article
Leila Roushangar and Jafar Soleimani Rad, 2007. Ultrastructural Alterations and Occurrence of Apoptosis in Developing
Follicles Exposed to Low Frequency Electromagnetic Field in Rat Ovary. Pakistan Journal of Biological Sciences, 10: 4413-4419.
DOI: 10.3923/pjbs.2007.4413.4419
URL: https://scialert.net/abstract/?doi=pjbs.2007.4413.4419
DOI: 10.3923/pjbs.2007.4413.4419
URL: https://scialert.net/abstract/?doi=pjbs.2007.4413.4419
INTRODUCTION
It is known that Electromagnetic Field (EMF), as an environmental factor, can produce functional and structural disorders. Progressive dependency of modern life to electrical devices has increased public concern about the potential effects of EMF on health. Humans are, indeed, continuously exposed to such harmful environmental factors produced by home appliances, diagnostic tools and industrial instruments with widely varying frequencies (Torregrossa, 2005; Genuis, 2007; Zymslony, 2007). It has been suggested that electric and magnetic fields at environmental levels may extend the lifetime of free radicals and result in DNA damage (Michael et al., 1999; Lai and Singh, 2004). EMFs could have a harmful effect on cellular metabolism by affecting biochemical reactions, behavior of charged molecules and ion channels, synthesis of macromolecules (Levin, 2003; Lai and Singh, 2004).
Some authors have reviewed risks of EMFs/Radiofrequency (RF) radiation on reproduction (Cheronff et al., 1992; Ahmed et al., 2002). Exposure to magnetic fields is reported to cause some adverse effects on female fertility (Huuskonen et al., 1993). These studies reviewed the potential effect of EMFs on infertility; implantation rate, number of living fetuses, sex ratio; miscarriages; premature births; Intrauterine Growth Retardation (IUGR), low birth weight, congenital malformations or genetic diseases, perinatal deaths and long-term effect like childhood cancer (Chiang et al., 1995; Feychting et al., 2005; Lahijani et al., 2007).
During the development of ovarian follicle in human, recruitment of a cohort of follicles occur, with only one destined to ovulate. The remainder of the cohort undergoes atresia by uncertain stimuli and mechanisms. It appears that growth factors and related peptides may are involved in this process (Gougeon, 1996; Wei and Giudice, 1997; Foghi et al., 1997). It is suggested that follicular atresia in the ovary results from apoptosis (Williams and Smith, 1993), in which cells die through activation of specific endogenous nucleases, resulting in certain morphological changes and DNA fragmentation (Billg et al., 1993). Morphological changes could be revealed by use of electron microscopy and the DNA breaks could be detected by applying terminal deoxyuridine nucleotide end-labeling assay (TUNEL). The role of apoptosis in follicular atresia has been extensively investigated in vivo and in vitro by molecular approaches in the rat (Chun et al., 1996), pig (Tilly et al., 1992), hamster (Bair et al., 1996), chicken (Tilly et al., 1991), rabbit (Kasuya, 1995) and cow (Jolly et al., 1994). The aims of the present study are) to study EMF-induced alterations in ovarian follicles at cellular level and to fined out whether exposure to EMF could increase the incidence of apoptosis in follicular cells and oocyte.
MATERIALS AND METHODS
Thirty adult female Wistar rats weighing 150-200 g were used in the present study. The animals were housed in plastic cages and kept in an air-conditioned room (temperature 24°C, humidity 45% and maintained on a 12 h light and 12 h dark cycle as previously reported (Flaws et al., 1994; Kao et al., 1999). The rats fed by laboratory chow and water ad libitum. Experiments were carried out in animal care unit and Drug Applied Research Center of Tabriz University of Medical Sciences, Iran.
The rats were divided into control and experimental groups. The rats in the experimental group are exposed to 3 milli Tesla EMF for 4 months, 4 h day-1. The EMF was produced by a device using 50 Hz alternative current. After the experimental period, for electron microscopic studies the tissue pieces of the ovaries dissected out and fixed in 2% glutaraldehyde in a 0.1 M phosphate buffer and post fixed in 1% OsO4. The specimens were then dehydrated through graded concentrations of ethanol and embedded in resin. One-micron semi-thin sections were stained with toluidine blue. Ultra thin-sections from selected blocks were stained with uranyl acetate and lead citrate and observed in a LEO 906 transmission electron microscope.
Morphometric studies were carried out on electron micrographs using common measurement tools.
For determination of DNA fragments in apoptotic cells the specimens were fixed in 10% neutral buffered formalin and DNA damage was detected by use of an in situ cell death detection kit (POD, Roche laboratories, Germany) and TUNEL staining was carried out according to manufacturer’s instructions. For assessment of apoptotic cells the stained specimens were viewed under bright field microscope and TUNEL positive cells were detected on the basis of their brownish color. Statical analysis was carried out using student t-test with the SPSS package to determine the significance of any differences seen in the parameters studied.
RESULTS
Control group: Healthy oocytes, in developing follicles, showed similar ultrastructural morphology regardless of the stage of development. Oocytes in follicles that developed to or beyond the primary stage acquired a zona pellucida, which encompassed and was tightly apposed to both the oocyte and the surrounding layer of granulosa cells (Fig. 1). The average width of zona pellucida in control group was 14.9±0.42 μm. In healthy follicles, the inner layer of granulosa cells, so called corona radiata, was composed of columnar cells and was in contact with the zona pellucida. Microvilli from the oocyte and granulosa cells were penetrated the zona pellucida (Fig. 1). The zona pellucida contained 9.8±0.56 microvilli sections per cm2. The cytoplasm of oocyte was sparsely populated with loosely clustered organelles. Mitochondria in oocytes appeared round or oval, rather dark and had a small number of identifiable shelf like crista. Numerous ribosome, poorly developed rough endoplasmic reticulum and some derbis-containing vesicles, with morphological features of secondary lysosomes, were also observed. A small groups of short, roughly parallel, linear structures, previously called lamellae, or cytoplasmic rays, were dispersed throughout the cytoplasm. Figure 2 shows the nucleus of an oocyte from control group. It demonstrates a regular and round euchromatic nucleus with surrounding nuclear envelope. The nucleus contain a well developed nucleoulus.
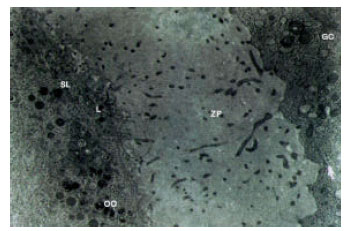 | |
| Fig. 1: | Transmission electron micrograph of a developing follicle from the control rat ovary demonstrating the Granulosa Cells (GC) and the Zona Pellucida (ZP) surrounding the Oocyte (OO). Penetrated microvilli from oocyte and coronal cells in the zona pellucida are obvious. In the cytoplasm of oocyte Lamella (L), a few Secondary Lysosomes (SL) and faintly stained mitochondria are seen. Junction between coronal cells is shown with arrow. X 3000 |
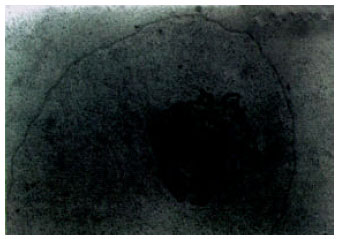 | |
| Fig. 2: | Transmission electron micrograph of the nucleus of a developing oocyte from the control rat ovary. Note a homogenous nuclear chromatin and a well developed nucleolus. X 10000 |
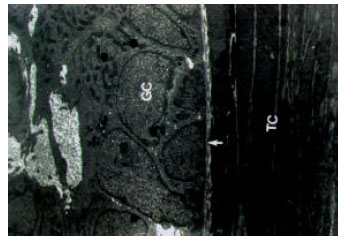 | |
| Fig. 3: | Transmission electron micrograph of developing follicle from the control rat ovary showing Granulosa Cells (GC), theca interna and Theca externa (TC), basement membrane (arrow). X 3000 |
In the developing follicles the granulosa layer composed of several cell layers and were enclosed by theca interna and externa (Fig. 3). The granulosa cells had euchromatic nuclei, several cytoplasmic organelles and were rest on a distinct basement membrane. There were some electrolucent spaces between granulosa cells. The theca interna was rich in cells and blood vessels, while the theca externa was rich in collagen fibers (Fig. 3).
EMF-exposed group: In the EMF-exposed rats, ultrastructural alterations in oocytes were apparent. That is; zona pellucida appeared more condensed and narrower than the control group.
 | |
| Fig. 4: | Transmission electron micrograph from an EMF-exposed developing follicle from a rat ovary. Note condensed granulosa cells (GC) which are lost contact with zona pellucida and adjacent cells. The Zona Pellucida (ZP) is narrow and devoid of microvilli from oocyte (O) and corona radiata cells. X 3000 |
The width of the zona pellucida was 10.8±0.25 μm which was significantly decreased (p<0.001) in comparison to the control group. Granulosa cells (coronal cells) were very condensed and lost contact with oocytes or neighboring cells. In comparison to control group, the microvilli of the oocytes were fewer in number and microvilli of the granulosa cells has lost or retracted from the zona pellucida. Therefore, the non microvilli sections in the zona pellucida were 4.13±0.83 cm-2, were significantly (p<0.001) lower than the control value. In the oocyte the organelles were more sparsely distributed throughout the cytoplasm and lamellae were increased. The rough endoplasmic reticulum were either lost or indistinguishable and mitochondria were smaller (Fig. 4). Additionally, ooplasm contained several lipid droplets (Fig. 4, 5). Granulosa cells (coronal cells) were very condensed and lost contact with oocytes or neighboring cells (Fig. 4). The nuclei of the oocytes in the EMF-exposed group appeared irregular in shape and the nucleoli were not as developed as in controls (Fig. 5). The basement membrane of granulosa cells was irregular and, in some follicles, disrupted. Some granulosa cells showed typical features of apoptosis such as nuclear condensation, formation of crescent like heterochromatin, changes in nuclear morphology, appearance of autophagic vacuoles and apoptotic bodies and separation of cells from the neighboring cells. Vesiculation of cytoplasm and disrupted mitochondria were also observed in the granulosa cells close to the basal lamina. The steroid secreting cells in the theca interna possessed condensed nuclei, dilated nuclear membrane,relatively large intracellular and intercellular spaces and several lipid droplets in the cytoplasm (Fig. 6).
| |
| Fig. 5: | Transmission electron micrograph of nucleus of an EMF-exposed developing oocyte from a rat ovary. Note irregularly outlined nucleus and a less developed nucleolus. X 10000 |
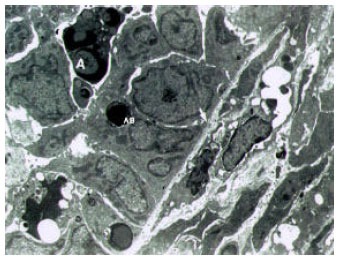 | |
| Fig. 6: | Transmission electron micrographs of an EMF-exposed developing follicle from a rat ovary. Apoptotic granulosa cells (A) apoptotic body in the cytoplasm of granulosa cell (AB), intracellular and intercellular spaces in the theca, basement membrane (arrow). X 3000 |
In this group, macrophages containing phagocytic vacuoles and apoptotic bodies were also frequently located near the wall of the blood vessels and among the stromal cells.
Histochemical studies for detection of apoptosis performed in sections from paraffin-embedded specimens using TUNEL technique. Brownish cells, representing TUNEL positive cells, were rare in the control group and detected in almost 10% of follicles. However, in EMF-exposed group, numerous TUNEL positive cells were present and could be detected in more than 70% of follicles. The cells were mainly located in the granulosa layer near the basal lamina and near the antrum. The TUNEL positive cells were also found in the granulosa layers of cystic follicles. Cystic follicles observed in both groups, but were frequent in EMF-exposed group.
DISCUSSION
The present study shows that EMF-exposure increases degenerative changes within the follicles. Present TEM has revealed that in EMF exposed group when compared with control group: The nuclei of oocytes were shrunken and had lost their regular shape. The oocytes had fewer microvilli and their cytoplasm contained a large amount of condensed lamellae. The zona pellucida, appeared narrower. The granulosa cells showed chromatin condensation and had several apoptotic bodies and autophagic vacuoles, corresponding to fat droplets in the control group. The alterations of oocytes and the presence of granulosa cells with condensed nuclei and their separation from neighboring cells constitutes the classical landmark of follicular atresia and is also characteristics of apoptosis (Tilly et al., 1992; Hurwitz and Adashi, 1991). It is known that the microvilli of oocyte and granulosa cells are in contact by gap junctions and are involved in oocyte nutrition (Devinc et al., 2006). The alterations produced by EMF could either be the result of initiation of apoptosis in the oocyte and follicular cells or would result in apoptosis in them.
EMF-induced alterations in the oocyte itself, could be considered as a pre apoptotic status of oocyte. Irregular morphology of nucleus could be an indication of changes in nuclear skeleton. Changes in cytoskeletal proteins and degradation of nuclear lamin is considered as a trigger of apoptosis cascade (Andrew et al., 1998). Probably our finding about morphological changes in the oocyte is very unique, in that, it correlate with apoptotic features. In this context, it is likely that, in rat ovary, EMF-exposure result in follicular atresia by inducing apoptosis in oocyte and granulosa cells.
Although the mechanisms underlying follicular atresia are not well known at this time, DNA damage, which can be initiated by oxidative free radicals, has been proposed as a possible mechanism that leads to the activation of the apoptotic cascade in atretic follicles (McLauchlan, 1980). In support of this hypothesis; it is shown that EMf has a pro-oxidant effect (Regoli et al., 2005) and it is proposed that the effect of EMF is mediated by production of free radicals (Bredt and Snyder, 1994). According to Peluso et al. (1977), graulosa cells in atretic follicles undergo nuclear condensation and cytoplasmic blebbing. Based on our sequential ultrastructural observations, these type of changes occurred in EMF-exposed granulosa cells and followed by appearance of apoptotic bodies, a characteristic of late apoptosis. Present findings well agrees with pro-oxidant effect of EMF (Regoli et al., 2005) and that, oxidants are well known apoptosis-inducing factors (Chandra et al., 2000; Jiang et al., 2002, 2005). The findings of the present study indicate that apoptosis of oocyte and granulosa cells are in part, responsible for initiation of follicular atresia. It also suggest that EMF, as an environmental factor, could disturb folliculogenesis by inducing apoptosis. Regarding the universal increase of infertility rate (Boivin et al., 2007), further studies are needed to clarify the relation between EMF exposure and infertility in human.
In the present study numerous autophagic and phagocytic vacuoles are also observed: The autophagic vacuoles are associated with the granulosa cells close to the basal lamina. As these vacuoles contain acid phosphatase activity, they are lysosomal in origin (Elfont, 1977) and in part responsible for the deterioration of the granulosa cell layer. The phagocytic vacuoles are observed in the thecal layers and interstitial tissue, associated particularly with the thecal cells and interstitial cells near the blood vessels. The deterioration of both granulosa and thecal cell layers is also enhanced by invasion of macrophages which occurs after many of the granulosa cells have undergone apoptosis. It has been proposed that apoptotic cells induce invasion of macrophages, possibly by releasing a chemotactic factor (Gaytan et al., 1998). In accordance with the aforementioned assumption, macrophages were frequently seen in EMF exposed group, it is known that macrophages have the capacity to produce oxidative products such as nitric oxide (Sugino et al., 1996), superoxide radicals and hydrogen peroxide (Byskov, 1974). It is also believed that macrophage-derived cytokines such as transforming growth factor, induces apoptosis in ovarian cells (Elfont et al., 1977).
It is proposed that macrophages scavenge degenerated cells in the ovary (Kerr et al., 1972; Leonard and Skeel, 1980). Thus, the apoptotic granulosa cells and thecal cells would be destroyed, leaving only fibroblasts and other connective tissue elements to represent the follicular wall, thereby transforming the follicle into a cystic follicle, a phenomenon which was frequently seen in EMF exposed group. These are in line with previous finding that the number of cystic follicles increases in ovaries from EMF-exposed rats (Soleimani Rad et al., 2003) and the result of the present study which shows the macrophages were frequent in the ovaries from EMF-exposed rats.
CONCLUSIONS
we have shown that EMF-exposure causes oocyte degeneration in the rat ovary by the mechanisms of apoptosis and cytotoxicity. This is evident in the EMF-exposed group with a large number of altered oocytes, apoptotic granulosa cells containing autophagic vacuoles and ruptured mitochondria, an increased number of macrophages and several lipid droplets in thecal cells. These findings well were correlated with the presence of TUNEL positive cells in the granulosa layer. Based on our TEM and histochemical studies, we have proposed an apoptosis inducing and cytotoxic effect for EMF in rat ovaries.
ACKNOWLEDGMENT
The authors are obliged to the Drug applied research center of the Tabriz University of Medical Sciences for their financial support. Thanks are also due to Mr. Daryush Mohammadnejad and Miss Ida aazami for their assistance in electron microscopy.
REFERENCES
- Elbetieha, A., M.A. Al-Akhras and H. Darmani, 2002. Long-term exposure of male and female mice to 50 Hz magnetic field: Effects on fertility. Bioelectromagnetics, 23: 168-172.
Direct Link - Bair, C.H., C.S. Chung, I.A. Vasilevskaya and W. Chang, 1996. Isolation and characterization of a Chinese hamster ovary mutant cell line with altered sensitivity to vaccinia virus killing. J. Virol., 70: 4655-4666.
Direct Link - Billg, H., I. Furuta and A.J. Hsueh, 1993. Estrogens inhibit and androgens enhance ovarian granulose cell apoptosis. Endocrinology, 133: 2204-2212.
Direct Link - Boivin, J., L. Bunting, J.A. Collins and K.G. Nygren, 2007. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod., 22: 1506-1512.
CrossRefDirect Link - Bredt, D.S. and S.H. Snyder, 1994. Nitric oxide: A physiologic messenger molecule. Ann. Rev. Biochem., 63: 175-195.
CrossRefDirect Link - Byskov, A.G.S., 1974. Cell kinetic studies of follicular atresia in the mouse ovary. J. Reprod Fertil., 37: 277-285.
CrossRefDirect Link - Chandra, J., A. Samali and S. Orrenius, 2000. Triggering and modulation of apoptosis by oxidative stress. Free Radical Biol. Med., 29: 323-333.
CrossRefPubMedDirect Link - Cheronff, N., J.M. Rogers and R. Kavet, 1992. A review of the literature on potential reproductive and developmental toxicity of electric and magnetic fields. Toxicology, 74: 91-126.
Direct Link - Chiang, H., R.Y. Wu, B.J. Shao, Y.D. Fu, G.D. Yao and D.J. Lu, 1995. Pulsed magnetic field from video display terminals enhances teratogenic effects of cytosine arabinoside in mice. Bioelectromagnetics, 16: 70-74.
Direct Link - Chun, S., K.M. Eisenhauer, S. Minami, H. Billig, E. Perlas and A.J.W. Hsueh, 1996. Hormonal regulation of apoptosis in early antral follicles: Follicle-stimulating hormones as a major survival factor. Endocrinology, 137: 1447-1456.
CrossRef - Devinc, P.J., C.M. Payn and M.K. Mccuskey, 2006. Ultrastructural evaluation of oocytes during atresia in rat ovarian follicles. Biol. Reprod., 63: 1245-1252.
Direct Link - Elfont, E.A., J.P. Roszka and M.J. Dimino, 1977. Cytochemical studies of acids phosphatase in ovarian follicles: A suggested role for lysosomes in steroidogenesis. Biol. Reprod., 17: 787-795.
Direct Link - Feychting, M., A. Ahlbom and L. Kheifets, 2005. EMF and health. Ann. Rev. Public Health, 26: 165-189.
Direct Link - Gaytan, F., C. Morales, C. Bellido and E. Aguilar, 1998. Ovarian follicle macrophages: Is Follicular atresia in the immature rat a macrophage-mediated event? Biol. Reprod., 58: 52-59.
Direct Link - Genuis, S.J., 2008. Fielding a current idea: Exploring the public health impact of electromagnetic radiation. Public Health, 122: 113-124.
CrossRefDirect Link - Gougeon, A., 1996. Regulation of ovarian follicular development in primates: Facts and hypotheses. Endo Rev., 17: 121-155.
Direct Link - Hurwitz, A. and E.Y. Adashi, 1992. Ovarian follicular atresia as an apoptotic process. Mol. Cell Endocrinol., 84: C19-C23.
Direct Link - Huuskonen, H., J. Juutilainen and H. Komulainen, 1993. Effects of low frequency magnetic fields on fetal development in rats. Bioelectromagnetics, 14: 205-213.
Direct Link - Jiang, S., S.E. Moriarty, H. Grossniklaus, K.C. Nelson, D.P. Jones and P. Sternberg Jr., 2002. Increased oxidant-induced apoptosis in cultured nondividing human retinal pigment epithelial cells. Invest. Ophthalmol. Vis. Sci., 43: 2546-2553.
Direct Link - Jiang, S., S.E. Moriarty-Craige, M. Orr, J. Cai, P. Sternberg Jr. and D.P. Jones, 2005. Oxidant-induced apoptosis in human retinal pigment epithelial cells: Dependence on extracellular redox state. Invest. Ophthalmol. Vis. Sci., 46: 1054-1061.
Direct Link - Jolly, P.D., T.J. Tisdall, D.A. Heath, S. Kun and K.P. Mcnatty, 1994. Apoptosis in bovine granulose cells in relation to steroid synthesis, cyclic adenosine 3, 5-monophosphate response to follicle-stimulating hormone and luteinizing hormone and follicular atresia. Biol. Reprod., 51: 934-944.
Direct Link - Kao, S.W., I.G. Sipes and P.B. Hoyer, 1999. Early effects of ovotoxicity induced by 4-vinylcyclohexene diepoxide in rats and mice. Reprod. Toxicol., 13: 67-75.
Direct Link - Kasuya, K., 1995. The process of apoptosis during the follicular epithelial cells in the rabbit ovary with special reference to involvement by macrophages. Arch. Histol. Cytol., 58: 257-264.
Direct Link - Kerr, J.F., A.H. Wyllie and A.R. Currie, 1972. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer, 26: 239-257.
PubMedDirect Link - Lai, H. and N.P. Singh, 2004. Magnetic field-induced DNA strand breaks in brain cells of the rat. Environ. Health Perspect., 112: 687-694.
Direct Link - Lahijani, M.S., S.E. Nojooshi and S.F. Siadat, 2007. Light and electron microscopic studies of effect of 50 Hz electromagnetic fields on preincubated chick embryo. Electromag. Biol. Med., 26: 83-98.
Direct Link - Michael, H., B. Repacholi and B. Greenebaum, 1999. Interaction of static and extremely low frequency electric and magnetic fields with living systems: Health effects and research needs. Bioelectromagnetics, 20: 133-160.
Direct Link - Regoli, F., S. Gorbi, N. Machella, S. Tedesco and M. Benedetti et al., 2005. Pro-oxidant effects of extremely low frequency electromagnetic fields in the land snail Helix aspersa. Free Radica. Biol. Med., 39: 1620-1628.
Direct Link - Sugino, N., K. Shimamura, H. Tamura, M. Ono, Y. Nakamura and K. Ogino, 1996. Progesterone inhibits superoxide radical production by mononuclear phagocytes in pseudopregnant rats. Endocrinology, 137: 749-754.
Direct Link - Tilly, J.L., K.I. Kowalski, A.L. Johnson and A.J.W. Hsueh, 1991. Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology, 129: 2799-2801.
Direct Link - Tilly, J.L., K.I. Kawalski, D.W. Schomberg and A.J. Hsueh, 1992. Apoptosis in atretic ovarian follicles is associated with selective decreases in messenger ribonucleic acid transcripts for gonadotropin receptors and cytochrome P450 aromatase. Endocrinology, 131: 1670-1676.
Direct Link - Torregrossa, M.V., 2005. Biological and health effects on electric and magnetic fields at extremely low frequencies. Ann. Ig., 17: 441-453.
Direct Link - Wei, Y.C. and L. Giudice, 1997. Programmed cell death in human ovary is a function of follicle and corpus luteum status. Clin. Endocrinol., 82: 3148-3155.
Direct Link - Williams, G.T. and C.A. Smith, 1993. Molecular regulation of apoptosis: Genetic controls on cell death. Cell, 74: 777-779.
Direct Link - Zmyslony, M., 2007. Biological mechanisms and health effects of emf in view of requirements of reports on the impact of various installations on the environment. Med. Pracy, 58: 27-36.
Direct Link