
Saima Malik
Department of Biotechnology, Faculty of Biological Sciences, Plant Cell and Tissue Culture Lab, Quaid-i-Azam University, Islamabad, Pakistan
Muhammad Zia
Department of Biotechnology, Faculty of Biological Sciences, Plant Cell and Tissue Culture Lab, Quaid-i-Azam University, Islamabad, Pakistan
Riaz-ur-Rehman
Department of Biotechnology, Faculty of Biological Sciences, Plant Cell and Tissue Culture Lab, Quaid-i-Azam University, Islamabad, Pakistan
M. Fayyaz Chaudhary
Department of Biotechnology, Faculty of Biological Sciences, Plant Cell and Tissue Culture Lab, Quaid-i-Azam University, Islamabad, Pakistan
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4118-4122







ABSTRACT
The study was conducted to analyze the effect of plant growth regulators on callogenesis and direct and indirect organogenesis of Memordica charantia. Callus cultures were induced from leaf, stem and cotyledonary explants of Momordica charantia, at different auxin and cytokinin concentrations either in single or in combination in MS medium. The best callogenic response was observed from all three explants (leaf, stem and cotyledon) on MS medium supplemented with 1.0 and 1.5 mg L‾1 BAP with 1.5 mg L‾1 NAA and 1.0 mg L‾1 2,4-D, respectively. The callus produced was hard, green and compact. These totipotent cells were failed to give rise shooting response when transferred to same or different growth regulator containing medium as second subculture. Indirect organogenesis response was very low or absent due to hardening of callus and habitutation. Best shooting was observed at 1.0 mg L‾1 BAP + 0.1 TDZ and 1.5 mg L‾1 BAP + 0.2 mg L‾1 NAA from shoot tip and cotyledonary node explants. While root formation was achieved when generated shoots were transferred to MS medium both full and half strength supplemented with different auxin concentrations.
PDF Abstract XML References Citation
How to cite this article
Saima Malik, Muhammad Zia, Riaz-ur-Rehman and M. Fayyaz Chaudhary, 2007. In vitro Plant Regeneration from Direct and Indirect Organogenesis of Memordica charantia. Pakistan Journal of Biological Sciences, 10: 4118-4122.
DOI: 10.3923/pjbs.2007.4118.4122
URL: https://scialert.net/abstract/?doi=pjbs.2007.4118.4122
DOI: 10.3923/pjbs.2007.4118.4122
URL: https://scialert.net/abstract/?doi=pjbs.2007.4118.4122
INTRODUCTION
The tissue culture technique is used for propagation, genotype modification, biomass production, germ plasm preservation and scientific investigations. These separate procedures have been collectively called tissue and organ culture (Ammirata, 1987) in vitro culture (Quak, 1997), micropropagation (Smith and Obeidy, 1991) and most recently biotechnology (Hartmann et al., 1997).
Bardford and Trewavas (1994) reported that plant growth regulators are involved in developmental transitions, such as flowering, embryogenesis, or dormancy and in real time responses to environmental conditions, such as adjustments in growth rates. Bud growth, shoot and root regeneration, root and stem hair formation, vascular tissue differentiation, abscission, ripening and flowering are examples of threshold phenomena that can be regulated by an inducing stimulus in a dose-dependent fashion.
Momordica is a genus of about 45 species of family cucurbitaceae, natives of tropical Africa and Southern Asia. Popularity of Momordica charantia in various systems of traditional medicines for several ailments (antidiabetic, contraceptive, jaundice, abdominal pain, kidney (stone), piles, pneumonia, fever etc.) focused the investigator’s attention on this plant (Grover and Yadav, 2004). A novel phytochemical in bitter melon has clinically demonstrated the ability to inhibit an enzyme named guanylate cyclase that is thought to be linked to the pathogenesis and replication of not only psoriasis, but leukemia and cancer as well (Takemoto, 1983). Other phytochemicals that have been documented with cytotoxic activity are a group of ribosome-inactivating proteins named alpha and beta-momorcharin, momordin and cucurbitacin B (Huang et al., 1996). The anticancerous and antileukemic activity of bitter melon against numerous cell lines including liver cancer, human leukemia, melanoma and solid sarcomas have also been documented (Zhu et al., 1990).
Manye et al. (2004) established an in vitro regeneration system of bitter melon (Momordica charantia L.). They proposed that during the period of inducing callus, the explants are not sensitive to the kinds and proportions of phytohormone. However, in the process of callus differentiating into adventitious bud, the kind, proportion and quantity of phytohormone and the type of callus made different result.
In vitro clonal propagation of Momordica charantia has been done by Agarwal and Kamal (2004) leading to shoot and root differentiation at different level of cytokinin and auxin in MS medium. Sultana and Bari (2003) observed direct shoot regeneration from nodal segments and shoot tips from in vitro grown seedlings of Momordica charantia L. when cultured on MS medium supplemented with different concentrations and combinations of cytokinins (BAP, Kin) and auxins (NAA, IAA). Thiruvengadam et al. (2006) also optimized a system for the somatic embryogenesis via embryogenic suspension cultures in bitter melon (Momordica charantia L.). Wang et al. (2001) developed a simple and efficient method of in vitro flowering of Momordica charantia from shoot tip explants by culturing on MS medium supplemented Sucrose, Fe2+ and different concentrations of cytokinin (BAP or Kin).
Due to the valuable factors of Momordica charantia its utilization as food and medicinal importance. Present study was conducted for micropropagation using different explants while preliminary work only describes shoot tip culture. Present study also describes a standard protocol to generate high number of plants and also standardization of tissue culture techniques to progress in the field of genetic transformation.
MATERIALS AND METHODS
Momordica charantia seeds were purchased from local market. The seed coat was removed and surface sterilized with 1% mercuric chloride and washed thoroughly with distilled autoclaved water under aseptic conditions. Seeds were germinated on plain agar medium containing 3% sucrose. After two weeks of germination, leaves, stems and cotyledons were excised at average size 2-3 cm and placed on pre-autoclaved Murashige and Skoog (1962) basal medium supplemented with different growth regulators for callus induction. While for direct organogenesis (shooting) cotyledonary node and shoot tip explants were placed on different concentrations/combinations of BAP, NAA and TDZ. Whereas for indirect organogenesis, primary calluses were transferred on regeneration medium after four weeks of callus initiation. All the cultures were kept in a cooled incubator with 16 h light cycle in every 24 h with temperature at 26±1°C. Shoots emerged were separated, callus was removed and planted again on full and half MS medium containing different concentrations of auxins for root initiation. Rooted plants were washed with distilled water and planted in soil and peat moss (3:1) under high moisture content for acclimatization. After one week, these plants were transferred to green house.
The experiments were entirely randomized with six replicates for each growth regulator sec‾1 concentration sec‾1. Statistical analyses were carried out by the ANOVA and Dunkens multiple test, at a 5% probability level.
RESULTS AND DISCUSSION
Callogenesis: Callogenic response from different explants varies depending upon morphological characteristics of explant and type and concentration of PGRs supplemented in MS medium (Table 1). 2,4-D, a sole PGR for callus induction produced maximum callus from all three explants (leaf, stem and cotyledons) but failed to produce callus at low level (0.1 and 1.0 mg L‾1). However, leaf explant showed maximum callus percentage and callogenic response then other two explants; stem and cotyledons, when tested on MS medium supplemented with BAP, NAA and Kin. The callus produced at these PGRs concentrations including 2,4-D, were of different texture and morphology depending upon nature of PGR. At 2,4-D all concentrations the calluses were brownish to yellowish green but compact and hard. While Thiruvengadam et al. (2006) stated that MS medium containing 1.0 mg L‾1 2,4-D approximately 90% of leaf explants of Momordica charantia L. gave rise to a well organized friable calli. At different concentrations of BAP and Kin green, compact and hard calluses produced. These calluses turned to be embryogenic under the stress of PGR. Berg et al. (1997) reported BAP as sole plant growth regulator, successfully preferred for good texture callus development. At different concentrations of NAA, the calluses produced were soft and yellowish green to green in color. These calluses also turned hard after four weeks. Best callogenic response was observed at different concentrations of BAP and NAA in combination but Callogenic response was low from all three explants. The calluses produced were soft and light green due to effect of cytokinin and auxin cumulatively. Nabi et al. (2002) found that for teasle gourd (Momordica dioica Roxb) callogenesis combination of 1.0 mg L‾1 BAP + 0.1 mg L‾1 NAA was most suitable that produced soft, light green and friable calli. At other combination and concentrations, texture, morphology and color of calluses varied depending on PGR supplemented in MS medium. However any explant failed to show callogenic response at hormone free MS medium.
Direct and indirect shooting response: Shooting response is dependent upon concentration of cytokinin supplemented in the medium. Cytokinin work as signaling molecules that activate totipotent cells of callus for shoot organogenesis where as in the case of direct organogenesis, these molecules activate preexisting machinery in the case of somatic cells (leaf, stem, cotyledon etc.) while in the case of shoot apex they stimulate the growth due to presence of meristemic cells at the tip of explant.
| Table 1: | Callogenic response of different explants of Memordica charantia at different concentrations of auxins and cytokinins |
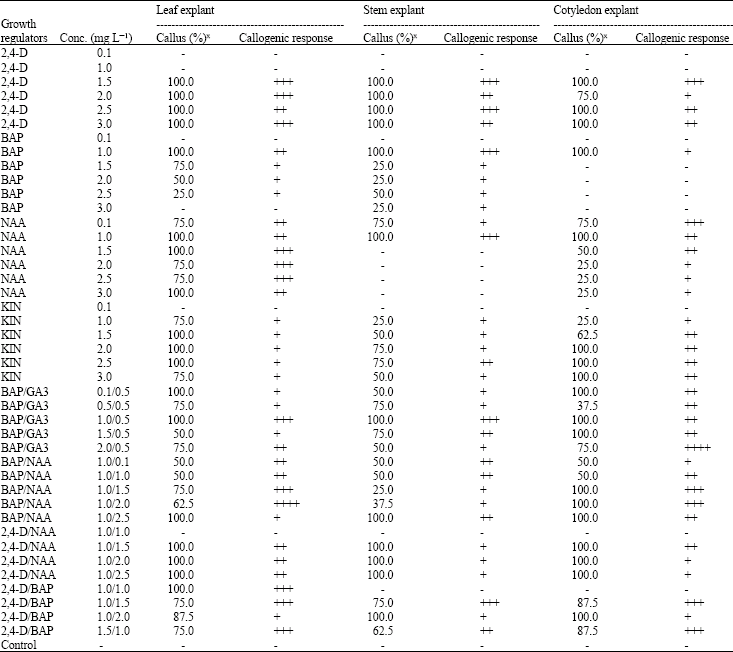 | |
These cytokinins may also turn the explant to produce multiple shooting response. Memordica callus did not produce any shoot at any concentration/combination tested (Table 2). At few combinations (BAP with TDZ or NAA), only leaves were formed that were lush green in color. At all concentrations tested the callus turned hard and embryogenic but failed to produce shoot. Manye et al. (2004) found that in the process of callus differentiating adventitious buds, the kind, proportion and quantity of phytohormone and the type of callus made different results in Momordica charantia L. They obtained adventitious buds from yellowish green callus on MS medium with BAP and Kin. However they observed very low differentiation from yellow or green calluses.
Shoot tip explant showed best shooting response (Table 2) at BAP with NAA (1.0+0.2 mg L‾1) supplemented in MS medium where average number of shoots per flask were 2.75±0.71 in which shoots attained maximum length of 1.74±0.69 cm. BAP with TDZ also exhibited good shooting response with average number of shoots 1.50±1.69 and length 0.800±0.899 cm. However at BAP alone shooting response was low. Same findings were of Pierik (1987) who stated that cytokinins are often used to stimulate growth and development, Kin and BAP being in common use. They usually promote cell division especially if added together with an auxin. At higher concentrations they can induce adventitious shoot formation by decreasing apical dominance and they retard aging. Cotyledonary node explants showed comparatively low response on same hormonal combinations giving maximum regeneration frequency 50.0% that was 100.0% in case of shoot tip explant (Fig. 1).
| Table 2: | Direct and indirect shooting response of Memordica charantia at different concentrations of plant growth regulators |
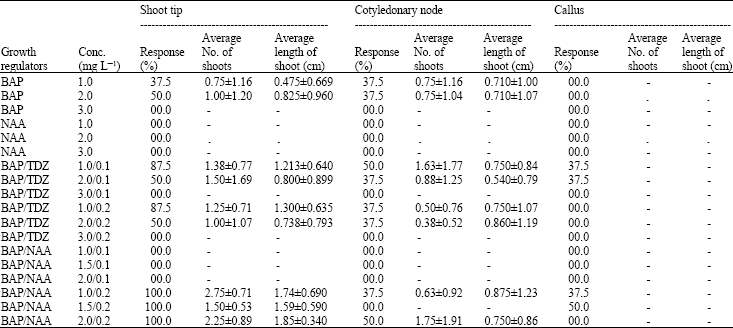 | |
 | |
| Fig. 1: | Shooting response from cotyledonary node explant and acclimatization of Memordica charantia |
From cotyledonary node explant highest shooting response at BAP/NAA combination gave average number of shoots 1.75±1.91 per flask with 0.75±0.86 cm maximum length. However Agarwal and Kamal (2004) observed shoot differentiation in Momordica charantia when alone BAP was used. They also obtained good results on NAA in combination with IBA. Sultana and Bari (2003) obtained similar results as that of present study. According to them best response towards multiple shoot regeneration was obtained from the nodal segments of Momordica charantia L. on MS medium supplemented with BAP and NAA. Hoque et al. (1995) also found that BAP and NAA as the best combination for adventitious multiple shoot formation in teasle gourd.
Best rooting response from generated shoots was observed at half strength of MS medium supplemented with 0.5 mg L‾1 NAA with 2.63±1.30 average number of roots giving 87.5% response and half strength MS without any growth regulator that gave 100% rooting response. While at full strength MS medium 0.1 mg L‾1 NAA gave maximum rooting response giving 1.38±1.06 average roots per flask. Same results were also observed by Agarwal and Kamal (2004) and Sultana and Bari (2003). Acclimatization percentage was 62.5%.
This study concludes that endogenous and exogenous level of growth regulators is also important for callogenesis and for differentiation. But habitutation property of callus hinders the organogenic response in Momordica charantia.
REFERENCES
- Agarwal, M. and R. Kamal, 2004. In vitro clonal propagation of Momordica charantia L. Indian J. Biotechnol., 3: 426-430.
Direct Link - Bardford, K.J. and A.J. Trewavas, 1994. Sensitivity threshold and variable time scale in plant hormone action. Plant Physiol., 105: 1029-1036.
Direct Link - Grover, J.K. and S.P. Yadav, 2004. Pharmacological actions and potential uses of Momordica charantia: A review. J. Ethnopharm., 93: 123-132.
CrossRefPubMedDirect Link - Manye, Y., Z. Maojun, Z. Yu, L. Liqong and C. Fang, 2004. Establishment of in vitro regeneration system of Bitter Melon (Momordica charantia L.). High Technol. Lett., 10: 44-48.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nabi, S.A., M.M. Rashid, M. Al-Amin and M.G. Rasul, 2002. Organogenesis in teasle gourd (Momordica dioica Roxb.). Plant Tissue Cult., 12: 173-180.
Direct Link - Smith, M.A.L. and A.A. Obeidy, 1991. Micropropagation of a mature male Kentucky coffee tree. Hortic. Sci., 26: 14-26.
Direct Link - Sultana, R.S. and M.A. Bari Miah, 2003. In vitro propagation of karalla (Momordica charantea Linn.) from nodal segment and shoot tip. J. Biological Sci., 3: 1134-1139.
CrossRefDirect Link - Takemoto, D.J., 1983. Purification and Characterization of a cytostatic factor with anti-viral activity from the bitter melon. Prep. Biochem., 13: 397-421.
PubMedDirect Link - Thiruvengadam, M., S. Varisai Mohamed, C.H. Yang and N. Jayabalan, 2006. Development of an embryogenic suspension culture of bitter melon (Momordica charantia L.). Sci. Hortic., 109: 123-129.
Direct Link - Wang, S., L. Tang and F. Chen, 2001. In vitro flowering of bitter melon. Plant Cell Rep., 20: 393-397.
Direct Link - Zhu, Z.J., Z.C. Zhong, Z.Y. Luo and Z.Y. Xiao, 1990. Studies on the active constituents of Momordica charantia. Yao Hsueh. Pao., 25: 898-903.
Direct Link