
Shokoofeh Hajihashemi
Iranian Academic Center for Education, Culture and Research, Alzahra University branch, Tehran, Iran
Khadijeh Kiarostami
Department of Plant Sciences, Alzahra University, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 1 | Page No.: 41-48







ABSTRACT
The effect of paclobutrazol (PBZ) treatment on salinity tolerance of wheat (Triticum aestivum), were investigated for two salt-tolerant and salt-sensitive cultivars. Salinity with PBZ treatment significantly reduced the plant height and length and area of sixth leaf in both cultivars. With increasing salinity, a gradually reduction was observed in roots length, fresh and dry weight of shoot and sixth leaf and relative water content of PBZ-applied plants in both cultivars. The greatest reduction was observed at 225 mM NaCl with 60 or 90 ppm PBZ. Salinity with PBZ treatment increased Na+ content in the sixth leaf and roots of both cultivars and the greatest increase was observed in salt-sensitive cultivar. In PBZ -treated plants, K+, P and N contents increased in line with elevating salinity in both cultivars, except at 90 ppm PBZ in salt-sensitive cultivar. Very similar effects of NaCl and PBZ treatment were observed for both cultivars regardless of their salinity susceptibility. The results suggest that PBZ treatment may be useful to improve the salt tolerance of wheat via reducing the negative effect of salinity on vegetative growth and the Na+ content and increasing the K+, P and N contents.
PDF Abstract XML References Citation
How to cite this article
Shokoofeh Hajihashemi and Khadijeh Kiarostami, 2007. Effects of Paclobutrazol and Salt Stress on Growth and Ionic Contents in Two Cultivars of Wheat. Pakistan Journal of Biological Sciences, 10: 41-48.
DOI: 10.3923/pjbs.2007.41.48
URL: https://scialert.net/abstract/?doi=pjbs.2007.41.48
DOI: 10.3923/pjbs.2007.41.48
URL: https://scialert.net/abstract/?doi=pjbs.2007.41.48
INTRODUCTION
Salt stress negatively impacts agricultural yield throughout the world, affecting production whether it is for subsistence or economic gain (Flowers, 2004). A considerable portion of irrigated land resources in arid and semi-arid regions of the world including Iran, where a large area has been rendered agriculturally unproductive due to high concentration of salt in the root zone. The United Nations Environment Program estimates that approximately 20% of agriculture land and 50% of cropland in world is salt-stressed (Flowers and YEO, 1995). According to Kamkar et al. (2004) 15% of Iran’s land is salt affected. Wheat is the source of almost 20% of total necessary calories for the world’s population (Perviaz et al., 2002). It is stable food of 43 countries including Iran, where it is grown on a large area.
Soil salinity causes significant reduction in plant productivity and consequent economic losses associated with reduced grain quality and yield of agricultural crops (Pitman and Läuchli, 2002). Salinity inhibits plant growth mainly by water deficit, ion toxicity and ion imbalance (Greenway and Munns, 1980). In wheat, genotypic variation in salt tolerance has been found to be associated with low rates of Na+ uptake and transport and high selectivity for K+ or Ca++ over Na+ (Schachtman and Munns, 1992). It is known that plants absorb the constituent ions of a saline substrate in varying degrees. This may result in harmful accumulations of a particular ions or decreasing the absorption of some essential nutrients beside the depressive effect on the activity of the endogenous growth hormones (Dawh et al., 1998). The presence of salt in soil solution decrease the osmotic potential of soil, thereby resulting in water stress and making in difficult for the plant to absorb water necessary for growth (El-Hendawy et al., 2005).
Attempts have been made to overcome the adverse effect of salinity, one of them is application of growth regulators. Paclobutrazol (PBZ) [(2RS, 3RS)-1-(4-chlorophenyl)-4, 4-dimethyl-2-(1, 2, 4-triazol)-pentan-3-ol] is a triazole plant growth regulator that consists of two enantiomers, namely 2R, 3R and 2S, 3S forms (Hedden and Graebe, 1985). PBZ blocks the biosynthesis of the active gibberellins (GA) and therefore decreases plant growth and development (Mehouchi et al., 1996). The triazole (such as PBZ) mediated stress protection is often explained in terms of hormonal changes such as an increase in cytokinins, a transient rise in abscisic acid (ABA) and a decrease in ethylene (Asar-Boamah and Fletcher, 1986; Fletcher and Hofstra, 1988; Mackay et al., 1990). Some researchers indicated that PBZ reduced the effect of salt stress on guava and grapes (Elaidy et al., 1992; Salama et al., 1992). The most pronounced effect of triazole on plants is a reduction in height (Fletcher and Hofstra, 1988). It was hypothesized that PBZ-treated plants had better quality of growth under salt stress than non-treated plants due to slower growth rate of former (Abou El-Khashab et al., 1997).
In spite of the increasing number of studies on PBZ-induced stress protection, a little is known about its effect on the physiological state of salt-stressed wheat plants. Thus, this report discusses the results of an experiment done to determine the effect of PBZ and salt stress and their interaction on vegetative growth, mineral content in the leaf and roots of two salt tolerance and salt sensitive cultivars of wheat.
MATERIALS AND METHODS
Plant culture and treatment: Seeds of two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat (Triticum aestivum) (Pustini, 2001) were provided from the Seed and Improvement Institute at Karaj, Iran. The experiment was conducted at Alzahra University, Tehran, Iran in November 2005. The seeds were cultured in 1.5 L pots, filled with vermiculite in a greenhouse with supplementary light to extend the photoperiod to 16 h per day. Air temperature ranged from 22 to 26°C during the day and 15 to 18°C during the night. Humidity ranged from 40 to 60%. Ten seeds were sown in each pot. The plants were irrigated every 4 days and fertilized every week with the standard Hoagland nutrient solution adjusted to pH. 6.0. Ten days after sowing, the seedlings were thinned to 4 similar plants per pot. At second leaf stage, the plants were sprayed with 0, 60 and 90 ppm of PBZ solution. PBZ treatment continued weekly during three consecutive weeks. Two days after the first PBZ treatment, the treated seedlings were irrigated with 0, 75, 150 and 225 mM of NaCl solution. Control plants were grown in Hoagland solution and sprayed with distilled water throughout the experiment. Salt treatment continued for one month and the plants were leached with tap water (after every three irrigations) to prevent salt accumulation. All measurements were performed two days after the last day of salt treatment.
Analysis of growth: Seedlings were randomly sampled at 45 days after planting, i.e., 6th to 7th leaf stage. Some growth parameters such as the plant height, roots length, length and area of sixth leaf, fresh and dry weight of shoot, roots and sixth leaf and relative water content (RWC) were measured from the collected seedlings. The plant height was measured from the base of the shoot to the liggule of the youngest fully extended leaf on the main stem. After the leaf area and the fresh weight of shoot, sixth leaf and roots were measured, the samples were washed with distilled water and dried in an oven at 70°C for 48 h and then the dry weight of samples were determined. Relative Water Content (RWC) was derived using the following equations:
RWC = FW-DW/DW |
Analysis of ions concentrations: Oven-dried samples of the sixth leaf and roots were ground into a fine powder. For determination of Na+, K+, P and N contents, 10 mg of ground dry material of the leaf and roots was digested by adding 4 mL HClO4 at 200°C until a colorless liquid achieved. After digestion, each sample brought up to a 25 mL final volume with distilled-deionized water. The Na+ and K+ contents were determined with a flame photometer (Cottenie, 1980; Isac and Robert, 1990; Waling et al., 1989). The samples were analyzed for the N content using Kjeldahl method (Cottenie, 1980; Isac and Robert, 1990; Jackson, 1997) and for the P content by spectrophotometric method (Champan and Pratt, 1961; Jackson, 1997).
Statistical analysis of data: A factorial experimental design with 2 genotypes, 4 salinity levels and 3 PBZ concentrations was arranged in a completely randomized block design with 4 replications. Data were analyzed by the Tukey and Duncan tests SPSS (version 12) statistical package to assess significant differences (at p≥5% level) among means.
RESULTS
Analysis of data showed that NaCl with PBZ treatment affected growth parameters. The effect of salt stress on the plant height, roots length, area and length of sixth leaf, fresh and dry weight of shoot and sixth leaf and RWC showed a similar tendency for both cultivars and decreased with increasing salinity, While the rate of reduction was difference among both cultivars (Fig. 1-5). For example, the greatest reduction in the plant height in salt-tolerant and salt-sensitive cultivars was by 62 and 51%, at 225 mM NaCl with 90 ppm PBZ (Fig. 1). In treated plants, the length of roots in cultivar Karchia was less than the control, whereas in salt-sensitive cultivar, the roots length showed no significant difference among the treated and control plants (Fig. 1). In spite of 150 and 225 mM NaCl with 90 ppm PBZ, which a markedly reduction was observed in the roots length of salt-sensitive cultivar.
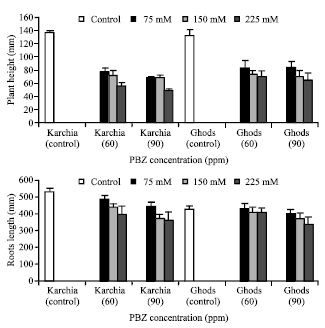 | |
| Fig. 1: | Effect of salt stress and PBZ treatment on the plant height and roots length in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
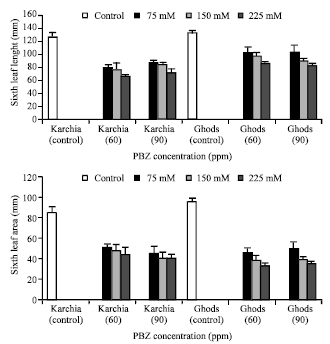 | |
| Fig. 2: | Effect of salt stress and PBZ treatment on the length and area of sixth leaf in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
The sixth leaf length of salt-tolerant cultivar at the same level of NaCl with PBZ was less than the salt-sensitive cultivar (Fig. 2). A highly significant reduction was achieved for the area of sixth leaf in both cultivars and the lowest reduction in the sixth leaf area in salt-tolerant and salt-sensitive was by 41% at 75 Mm NaCl with 60 ppm PBZ and 47% at 75 mM NaCl with 90 ppm PBZ, respectively (Fig. 2).
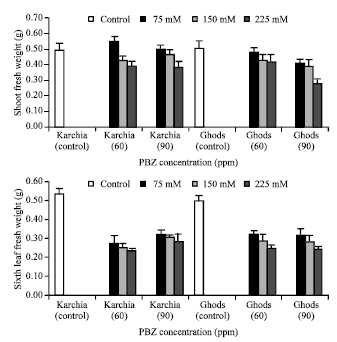 | |
| Fig. 3: | Effect of salt stress and PBZ treatment on the fresh weight of shoot and sixth leaf in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
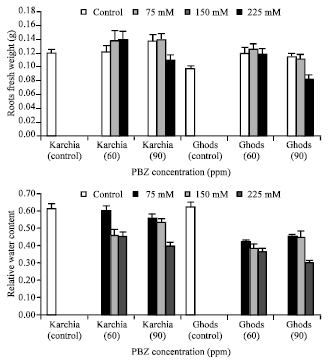 | |
| Fig. 4: | Effect of salt stress and PBZ treatment on the fresh weight of roots and relative water content (RWC) in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
The fresh weight of shoot in treated plants was less than the un-treated plants in both cultivars, except at 75 mM NaCl with 60 and 90 ppm PBZ (Fig. 3). In plants subjected to the salt with PBZ treatment, the fresh weight of sixth leaf significantly reduced in both cultivars and the greatest reduction was observed at the highest level of salinity (Fig. 3). In the control plant, the fresh weight of roots was less than the treated plants of both cultivars, but a noticeably reduction was observed at 225 mM NaCl with 90 ppm PBZ (Fig. 4).
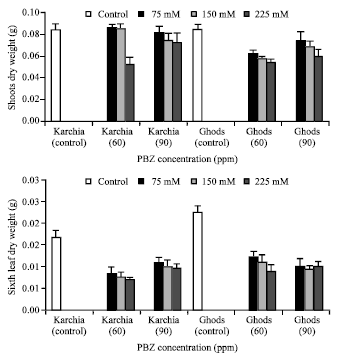 | |
| Fig. 5: | Effect of salt stress and PBZ treatment on the dry weight of shoot and sixth leaf in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
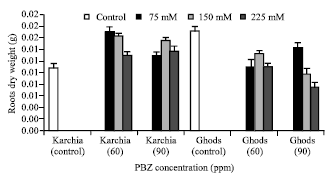 | |
| Fig. 6: | Effect of salt stress and PBZ treatment on the dry weight of roots in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
The negative effect of treatment on RWC in salt-sensitive cultivar was more than salt-tolerant cultivar because the average value of reduction in salt-tolerant cultivar was less than salt-sensitive cultivar (Fig. 4).
Under NaCl with PBZ treatment condition, the dry weight of shoot reduced in both cultivars, whereas a slightly increase was observed at 75 and 150 mM NaCl with 60 ppm PBZ in cultivar Ghods (Fig. 5). In treated plants, the sixth leaf dry weight markedly decreased and the greatest reduction in cultivars Karchia and Ghods was observed at 225 mM NaCl with 60 ppm PBZ, by 45 and 51%, respectively (Fig. 5).
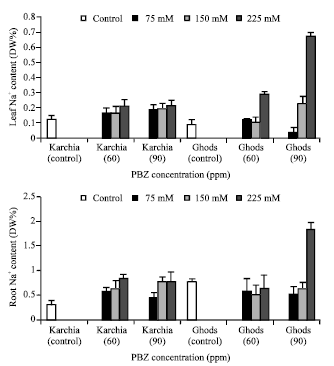 | |
| Fig. 7: | Effect of salt stress and PBZ treatment on the Na+ content of the sixth leaf and roots in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
So the reduction in sixth leaf dry weight of cultivar Ghods was significantly more than cultivar Karchia. The dry weight of roots in salt-sensitive cultivar was visibly more than salt-tolerant cultivar (Fig. 6). NaCl with PBZ treatment significantly increased the roots dry weight in salt-tolerant cultivar, while a significant reduction was observed in salt-sensitive cultivar.
The NaCl with PBZ treatment maintained the Na+ content in the sixth leaf and roots of cultivar Krchia (Fig. 7). The content of Na+ in the sixth leaf and roots of treated plants reduced in cultivar Karchiae, except at 150 and 225 mM NaCl with 90 ppm PBZ in the sixth leaf and 225 mM NaCl with 90 ppm PBZ in the roots, where a highly significant increase was observed. In PBZ-applied plants, the K+ content in the sixth leaf of salt-tolerant cultivar significantly increased with increasing salinity and a significant increase was observed at 225 mM NaCl with 60 ppm PBZ (Fig. 8). The measurements of K+ content in the roots showed no significant difference among the treated and control plants, except at 225 mM NaCl with 60 ppm PBZ. The amount of K+ in the sixth leaf and roots of cultivar Ghods significantly decreased in treated plants. The P content of sixth leaf and roots of PBZ-treated plants increased in line with elevating salinity in both cultivars, in spite of 225 mM NaCl with 90 ppm PBZ in salt-sensitive cultivar (Fig. 9).
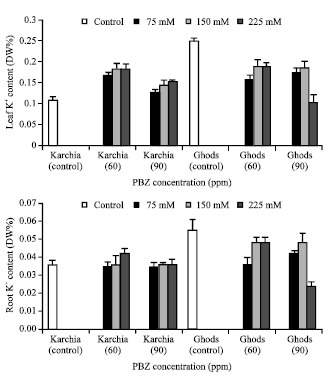 | |
| Fig. 8: | Effect of salt stress and PBZ treatment on the K+ content of the sixth leaf and roots in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
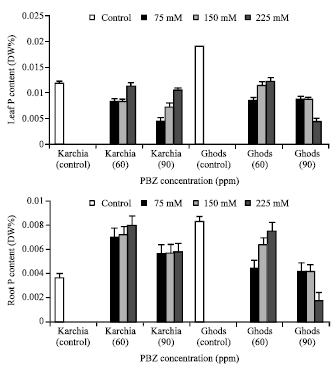 | |
| Fig. 9: | Effect of salt stress and PBZ treatment on the P content of the sixth leaf and roots in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
The amount of P in the sixth leaf and roots of treated plants was less than the control plants in both cultivars, but the P content in the roots of cultivar Karchia significantly was more than the control. Under treatment condition, the N content in the sixth leaf of salt tolerant cultivar significantly was more than the control, against the salt-sensitive cultivar (Fig. 10).
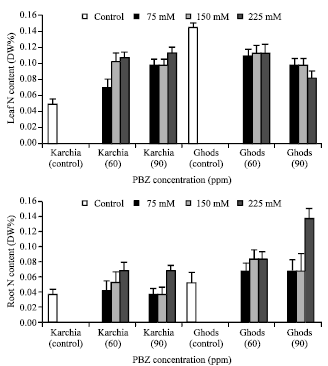 | |
| Fig. 10: | Effect of salt stress and PBZ treatment on the N content of the sixth leaf and roots in two salt-tolerant (Karchia) and salt-sensitive (Ghods) cultivars of wheat |
In both cultivars, the roots of treated plants accumulated more N content than the control, except at 75 and 150 mM NaCl with 90 ppm PBZ in cultivar Ghods.
DISCUSSION
In saline environment where salt are present in higher concentration, plant growth is affected negatively in various ways such as osmotic effect, specific ion effect and nutritional imbalance, probably all occurring simultaneously (Flowers et al., 1991). Initial growth inhibition in saline environment is induced by the decreased water potential of rooting medium due to higher salt concentration (Munns et al., 1995). The growth of both cultivars in terms of the plant height, roots length, sixth leaf length and area, fresh and dry weight of shoot, roots and sixth leaf and RWC was affected by PBZ treatment with salt stress. The decrease in shoot and roots yield of all cultivars with the addition of salt could be due to reduction of physiology availability of water with the increase in solute suction from saline media or accumulation of toxic ions in plants (Perviaz et al., 2002). With increasing NaCl concentration from 75 to 225 mM, a gradually reduction was observed in the plant height, roots length and length, area, fresh and dry weight of sixth leaf in both cultivars, while it was predicted that salinity cause a significant reduction in these parameters only in salt-sensitive cultivar. It is hypothesized that PBZ treatment moderated the negative effect of salinity on growth parameters in salt-sensitive cultivar. As result showed, PBZ significantly decreased the plant height, leaf length and area in both cultivars. Reduced height in a consequence of Triazole-induced GA inhibition, exemplified by reduced internodal elongation (Fletcher et al., 2000). PBZ reduces cell number and length, so PBZ-treated plants characteristically have smaller leaf. Measurements of the roots length indicated that PBZ improve the roots length of cultivar Ghods in salt-stressed plants. The increased root diameter has been associated with larger parenchyma cells and the promotion of radial cell expansion (Fletcher et al., 2000). Increasing NaCl concentration slowed down water uptake by plants, thereby the RWC decreased which followed by reduction of the fresh weight in both cultivars, especially in salt-sensitive cultivar. Inhibition of growth and decrease in water content induced by water stress has been universally observed even in tolerant plants. Although growth is a visible indicator of plant performance under stress, it is to result from the sum of the adaptive mechanism that is adapted by a given species. Therefore, it would predict that PBZ can induce adaptive mechanisms in wheat. It is hypothesized that, the fresh weight of shoot and roots increased in PBZ-treated plants because the water content enhanced. The leaves of PBZ-treated plants were significantly smaller, therefore the fresh and dry weight of leaf decreased in comparing with unsprayed plants. Decreased dry weight of shoot correlated with decreased plant height and smaller leaf in PBZ-treated plants. PBZ reduced the harmful effect of salinity on plants by a significant increase in the fresh weight of roots, comparing to un-treated plants. Protection of PBZ-treated plants from salinity was associated with smaller leaf for loosing less water which improved RWC in salt-stressed plants. Inhibition of GA by PBZ followed by a shift in hormonal balance including reduction in ethylene and increase in cytokinins and ABA are crucial changes associated with PBZ induced stress tolerant in plants (Fletcher et al., 2000).
A secondary effect of high concentrations of Na+ and Cl¯ in the root medium is the suppression of uptake of essential nutrients such as K+, Ca++ and NO3¯, etc (Perviaz et al., 2002). Na+ competes with K+ for uptake through common transports systems and dose this effectively since the Na+ is saline environments is usually considerably greater than K+ (Maathuis et al., 1996; Rains and Epstein, 1967). Salt-tolerant plant have low Na+ content in the leaf and stem, suggesting that these genotypes had a better ability to exclude harmful ions from the shoot, which in turn to contributed to their salt-tolerant (Lindsay et al., 2004). The Na+, K+, P and N contents in investigated wheat tissues were strongly affected by salinity with PBZ treatment. As expected, the Na+ content in the sixth leaf and roots increased with increasing salinity and the most amounts was observed at 225 mM NaCl. In Ghods cultivar no significant difference was observed among 60 and 90 ppm PBZ, whereas at the highest level of salinity in salt-sensitive, the Na+ content at 90 ppm PBZ was significantly more than 60 ppm PBZ. The treated plants of both cultivars showed a different tendency for accumulating Na+ content, so the Na+ content is a suitable indicator for recognizing salt tolerance cultivar. It is concluded that PBZ treatment inhibited elevation the Na+ content in salt-sensitive cultivar. Sodium exclusion from the transpiration stream reaching the leaf is controlled at three stages: (1) by selectivity of root cells taking up cations from the soil solution, (2) by selectivity in the loading of cations into the xylem vessels in the roots and (3) by removal of sodium from the xylem in the upper part of roots and the lower part of the shoot (Munns et al., 2002; Tester and Devenport, 2003). In saline soils, salinity not only causes high Na+ accumulation in plants, it also influences the uptake of essential nutrients such as K+ through effect of ion selectivity. The reduced K+ in plants, in turn, affect the integrity and functioning of cell membranes under saline conditions, which has been suggested to be an important selection citration for salt-tolerant (El-Hendawy et al., 2005). The increases in K+, N and P in the sixth leaf and roots of cultivar Ghods were observed in conjunction with elevating salinity in response to PBZ treatment. Studied showed that concentration of K+ in salt stressed plants were decreased with increasing salinity, an observation was also made by (Sepehr et al., 2003). PBZ treatment moderated the effect of stress on essential nutrients. It would be predicted that salinized plants accumulated more K+, P and N contents to degrade stress damages. PBZ treatment increased K+, P and N content in line with elevating salt concentration, against the effect of salinity alone. In salt-sensitive cultivar, the effect of 60 ppm PBZ on the amount of K+, P and N is more than 90 ppm PBZ and at 60 ppm PBZ with 150 and 225 mM NaCl the plants accumulated more K+ and P than 90 ppm PBZ with the same salinity level, especially at 225 mM NaCl.
It is noticeable that, the effect of salt stress on both cultivars was similar regardless of the salinity tolerance. So it is reasonable to conclude that PBZ is effective in reducing stress that occurs regularly with salt. Reduction in excessive vegetative growth and the Na+ content and concomitant increase in the K+, P and N contents in response to PBZ moderated the harmful effect salinity on both cultivars especially in salt-sensitive cultivar. PBZ has the potential to increase the productivity wheat in the saline areas where high salinity limits its introduction and cultivation. Our results suggest that 60 ppm PBZ is more effective than 90 ppm PBZ in moderating the negative effect of salt stress. Further research and field experiment are needed to indicate the addition pathways that improve salt-tolerant in PBZ-treated plants.
REFERENCES
- Abou El-Khashab, A.M., A.F. El-Sammak, A.A. Elaidy and M.I. Salama, 1997. Paclobutrazol reduces some negative effect of salt stress in peach. J. Am. Soc. Hortic. Sci., 122: 43-46.
Direct Link - El-Hendawy, S.E., Y. Hu and U. Schmidhalter, 2005. Growth, ion content, gas exchange and water relations of wheat genotypes differing in salt tolerances. Aust. J. Agric. Res., 56: 123-134.
Direct Link - Sepehr, M.F., M. Ghorbanli and R.A.K. Nejad, 2003. Effects of cadmium and salinity on growth, photosynthesis and ionic contents of Zea mays. Asian J. Plant Sci., 2: 196-201.
CrossRefDirect Link - Fletcher, R.A., A. Gilley, N. Sankhla and T.D. Davis, 2000. Triazoles as plant growth regulators and stress protectants. Hortic. Rev., 24: 55-138.
Direct Link - Flowers, T.J. and A.R. Yeo, 1995. Breeding for salinity resistance in crop plants: Where next? Aust. J. Plant Physiol., 22: 875-884.
CrossRefDirect Link - Mackay, C., J. Hall, G. Hofstra and R. Fletcher, 1990. Uniconazole-induced changes abscisic acid, total amino acids and proline in Phaseolus vulgaris. Pest. Biochem. Physiol., 37: 74-82.
Direct Link - Mehouchi, J., F.R. Tadeo, S. Zaragoza, E. Promo-Millo and M. Talon, 1996. Effect of gibberellic acid and paclobutrazol on growth and carbohydrate accumulation in shoots and roots of citrus rootstock seedlings. J. Hortic. Sci., 71: 747-784.
Direct Link - Munns, R., D.P. Schachtman and A.G. Condon, 1995. The significance of a two-phase growth response to salinity in wheat and barley. Aust. J. Plant Physiol., 22: 561-569.
CrossRefDirect Link - Salama, M.I., A.A. Elaidy, A. El-Sammak and A.M. Abou-El-Khashab, 1992. Leaf pigments and nutrient elements content of Roumi Red grape nursling as affected by salinity and some growth regulators. Tanta J. Agric. Res., 18: 382-391.
Direct Link - Schachtman, D.P. and R. Munns, 1992. Sodium accumulation in leaf of Triticum species that differ in salt-tolerant. Aust. J. Plant Physiol., 19: 331-340.
Direct Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link