
Ali Mojtahedi
Department of Medical Microbiology, Faculty of Medicine, Guilan University Complex, Rasht-Tehran Road, Rasht,3477, Iran
Rasoul Salehi
Genetics and Molecular Biology,
Farahtaj Navabakbar
Department of Medical Microbiology,
Hasan Tamizifar
Department of Medical Microbiology,
Hamid tavakoli
Internal Medicine (gastroenterology), School of Medicine,
Isfahan University of Medical Sciences, Isfahan, 81744-176, Islamic Republic of Iran
Vincent Duronio
Department of Experimental Medicine, School of Medicine, British Columbia University, Vancouver, Canada
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4097-4102







ABSTRACT
Helicobacter pylori is one of the most common pathogens affecting humans and is the major environmental factor in the development of gastric cancer increasing from 4 to 6 folds the risk of its development. Variations in cancer risk among H. pylori infected individuals may correlate to difference in H. pylori strains, variable host characteristics and specific interactions between host and microbial determinants. To determine the effect of different strains of H. pylori on cellular apoptosis this study was designed an in vitro model using AGS and HEF cell lines. After specified time intervals total cell proteins was extracted and subjected to SDS-PAGE and immunoblotting using anti poly ADP-ribose polymerase (PARP) antibody. Decrease in densitometric value of PARP was indicative of higher level of apoptosis. The ability of apoptosis induction in AGS and HEF cell lines by wild type (cagA+/vacA+), cagA¯/vacA+, vacA¯/cagA+ and double negative (cagA¯/vacA¯) strains were significantly different. The assessed apoptosis in AGS cell line co-cultured with wild type strain was 3.22±0.2 in 24 h, 2.8±0.1 in 48 and 2.1±0.09 in 72 h of incubation time. Similar assessment with cagA¯/vacA+ strains in AGS cells was 4.17±1.49 in 24 h, 3.32±0.45 in 48 h and 2.32±0.61 in 72 h incubation. A variation in apoptotic potential between the H. pylori strains on two cells (AGS and HEF) was observed. Based on present results, it is concluded that H. pylori strains as well as target cell types are important in pathogenesis and induction of apoptosis during a specified time interval.
PDF Abstract XML References Citation
How to cite this article
Ali Mojtahedi, Rasoul Salehi, Farahtaj Navabakbar, Hasan Tamizifar, Hamid tavakoli and Vincent Duronio, 2007. Evaluation of Apoptosis Induction Using PARP Cleavage on Gastric Adenocarcinoma and Fibroblast Cell Lines by Different Strains of Helicobacter pylori. Pakistan Journal of Biological Sciences, 10: 4097-4102.
DOI: 10.3923/pjbs.2007.4097.4102
URL: https://scialert.net/abstract/?doi=pjbs.2007.4097.4102
DOI: 10.3923/pjbs.2007.4097.4102
URL: https://scialert.net/abstract/?doi=pjbs.2007.4097.4102
INTRODUCTION
Helicobacter pylori infects over half of the world’s population and causes a wide range of diseases, including gastritis, peptic ulcer and two forms of gastric cancer. (Guillemin, 2002).
However, only a fraction of individuals who carry H. pylori ever develop gastric neoplasia. Variations in cancer risk among H. pylori-infected individuals may be related to differences among H. pylori strains, variable host characteristics, environmental influences and/or specific interactions between host and microbial determinants (Cover et al., 2003). Different hypothesis put forward to explain the underlying mechanism of carcinogenesis by H. pylori. It is generally believed that this carcinogenic activity is due to disturbance between epithelial apoptosis and proliferation. However, there are various reports on effect of different strains of H. pylori on apoptosis induction in different target cells.
The cagA (cytotoxic associated gene A) and vacA (vacuolating cytotoxin A) genes present in the wild type H. pylori are strongly related to the bacterial ability to induce adverse changes in stomach epithelium (Ashktorab et al., 2004; Cover et al., 2003; Hua et al., 2000; Jones et al., 1999; Kuck et al., 2001). These changes are initiative points in carcinogenic pathway induced by H. pylori. The mutant strains deprived one or both of these virulence related genes exhibit reduced pathogenicity compared to wild type H. pylori.
Although apoptosis may be a natural physiological occurrence, excessive apoptosis results in tissue damage. Alternatively, apoptosis may be viewed as the response to hyper proliferation in an attempt to reduce tissue growth. For instance; Peek et al. (1999) have reported H. pylori cag+ strains enhance gastric epithelial cell proliferation and attenuate apoptosis in vivo, which may partially explain the increased risk of gastric cancer (Peek et al., 1999). Also Anti et al. (1998) have reported that there is no evidence that H. pylori or its cytotoxic products have any mutagenic effects. Nevertheless, this infection is associated with profound changes in the pattern of epithelial cell turnover in gastric glands, though the importance of these changes in gastric carcinogenesis is still controversial. H. pylori infection increases cell proliferation and alters the distribution of cycling cells within these glands (Anti et al., 1998). On the other hand, it was hypothesized that H. pylori may induce hyper-proliferation through increasing apoptosis (Moss et al., 2001). Whether apoptosis is the primary or secondary event, is not clear, but extrapolation from the data derived in cell culture would suggest that apoptosis is the initial epithelial cell response. Thus, the induction of excessive apoptosis by H. pylori could induce a secondary hyper proliferative response in an attempt by the mucosa to maintain cell mass (Shirin and Moss, 1998).
Once hyper proliferation is established, then perhaps the increased rate of cell cycling predisposes gastric epithelial cells to genotoxic damage and an altruistic cell death. If this altruistic pathway fails, then unrestrained tissue growth may result (Shirin and Moss, 1998). This process of programmed cell death is characterized by marked changes in cell morphology and the cleavage of several housekeeping proteins including poly (ADP-ribose) polymerase (PARP).
Considering these controversial reports on association of different strains of H. pylori to the carcinogenic process and their role in induction of apoptosis, evaluation of the process in a well designed study was appeared to be highly needed.
Apoptosis was determined in various time intervals when H. pylori and cells were co-cultured together. PARP cleavage was used as criteria for occurrence of apoptosis (Shibayama et al., 2001).
MATERIALS AND METHODS
Cell culture: AGS (gastric adenocarcinoma, ATCC CRL 1739) and HEF (Human embryonic fibroblast ATCC CRL-7093) cells were purchased from American Type Culture Collection (USA). AGS cells were cultured in 6 cm plates in Ham's F-12 medium (Gibco, USA) supplemented with 10% heat inactivated fetal calf serum without antibiotics. Cells were incubated in 5% CO2 at 37°C in humidified air. HEF cells were cultured in DMEM supplemented with 10% heat-inactivated fetal calf serum, L-glutamine and sodium pyrovate without antibiotics. The incubation condition was same as AGS cell line. The concentration of both cells was 1χ106 cells in each plate. The cells were left to adhere for 24 h.
Bacterial strains: Four strains of H. pylori were used in this study. One of them was cagA positive and cytotoxin producing H. pylori strain (wild type = cagA+/vacA+) and three strains were cagA&oline/vacA+, vacA&oline/cagA+ and double negative (cagA&oline/vacA&oline), respectively. First three strains were generously provided by Dr. Victoria S. Conlin (Physiology Department, UBC, Canada).
H. pylori strains were grown for 3-4 days at 37°C in Columbia agar (Difco, USA) under microaerophilic conditions obtained by using campy pak plus (B.D., USA) and an anaerobic jar supplemented with 5% defibrinated horse blood (Oxoid, USA), 10% heat-inactivated fetal calf serum, Trimethoprim, cefsulodine and Vancomycin (Sigma, USA) (Conlin et al., 2004). The cells then harvested and resuspended in Brucella broth (Difco, USA) supplemented with 10% heat-inactivated fetal calf serum, IsovitalX (BBL, USA) and kanamycin (for mutants).
H. pylori strains were grown 5-7 days in falcon tubes with shaking at 200 rpm, at 37°C under microaerophlic conditions (Conlin et al., 2004).
Co-culture experiment: For co-culture experiment, H. pylori strains were added to cells at ratio of 100:1, based on previous reports (Chen et al., 1997). For this purpose, the bacterial densities were adjusted by the optical density (OD) measurement at 600 nm. Calculation of bacterial concentration made on the basis of 1 OD600 = 108 colony forming units cfu mL&oline1 (Peek, 2002).
Bacteria were harvested and resuspended with Ham's F-12 and DMEM separately for AGS and HEF, respectively.
AGS and HEF cells were washed 3 times with sterile PBS and different strains of bacteria were added separately to AGS and HEF cells and incubated at 37°C in 5% CO2 for 0, 24, 48 and 72 h. Cells without bacteria were grown for control.
SDS-PAGE and Immunoblotting analysis: Co-cultured cells with bacteria and their controls were lysed in solubilization buffer (50 mM Tris-HCl (pH 7.7), 1% Triton X-100, 10% Glycerol, 100 mM NaCl, 2.5 mM EDTA, 10 mM NaF (Na Flouride), 0.2 mM NaV (Na orthovanadate), 1 mM NaM (Na Molybdate), 40 μg mL&oline1 PMSF (Phenylmethylsulphonylfluoride), 1 μM pepstatin, 0.5 μg mL&oline1 leupeptin and 10 μg mL&oline1 soybean trypsin inhibitor) (Peek, 2002).
Total cell protein extracts were normalized for concentration by the BCA (Bicinchoninic acid) assay and 50 μg of protein separated by SDS-PAGE and transferred to nitrocellulose membrane. Membranes were incubated overnight at 4°C with mouse anti-PARP monoclonal antibody (B.D. Pharmingen, USA).
Primary antibody was detected using anti mouse horse radish peroxidase-conjugated secondary antibody for 1 h and visualized by the ECL (Enhanced chimicoluminiscent) detection system (Amersham Corp, UK) according to the manufacturer's instructions.
Statistics: Results are expressed as means±standard errors (SD). t-test, one-way analysis of variance (ANOVA) was used, followed by post hoc comparisons, general linear multivariate and repeated measure.
RESULTS
To determine whether infection with different strains of H. pylori alone could stimulate apoptosis process, AGS and HEF cell lines were incubated with four different strains of H. pylori in the ratio of 1:100 (cell: bacteria) for 24, 48 and 72 h.
All experiments were performed 3-5 times and the mean values were taken for comparison.
Quantitation of apoptotic induction of different strains of H. pylori on AGS and HEF cells was determined by rate of PARP cleavage using UN-SCAN-IT software. Decreased in densitometric value of PARP indicates its elevated cleavage and hence, pronounced apoptotic activity.
Figure 1, shows a representative western blot for PARP cleavage results.
The apoptotic activity of wild type H. pylori on AGS cells was calculated 3.22±0.2 in 24 h, 2.8±0.1 in 48 h and 2.1±0.09 in 72 h, respectively.
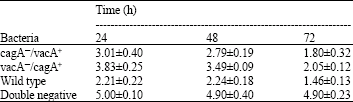
Similarly on HEF cell line 2.21±0.22 in 24, 2.24±0.18 in 48 and 1.46±0.13 in 72 h obtained when H. pylori was co-cultured with them.
To compare between two different types of cell in different continues time, we conducted GML (general linear model). GML showed that apoptotic potential influence between the two cell lines was statistically significant (p<0.05).
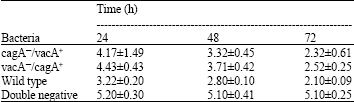
The apoptosis induced by cagA&oline/vacA+ strains of H. pylori in AGS cells was 4.17±1.49 for 24 h, 3.32±0.45 for 48 h and 2.32±0.61 for 72 h and in HEF cells was 3.01±0.4 for 24 h, 2.79±0.19 for 48 h and 1.8±0.32 for 72 h.
Analysis with independent t-test showed that apoptotic potential influence between the two cell lines does not statistically significant (p>0.05).
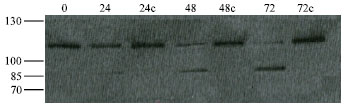 | |
| Fig. 1: | Representative western blot for PARP cleavage results. The above western blot analysis of PARP cleavage is shown for AGS cell line treated with WT strain of Helicobacter pylori in different times. The columns 0 to 72c show time 0 to 72 h. 24c, 48c and 72c are for controls. The 116 kDa intact form of PARP is seen in both untreated and Helicobacter pylori treated cell lysates. However, the 85 kDa PARP cleaved fragment is seen only in the treated cell line |
| Table 1: | Effect of different strains of H. pylori on induction of apoptosis using AGS gastric adenocarcinoma cell line |
 | |
| Table 2: | Effect of different strains of H. pylori on induction of apoptosis using HEF fibroblast cell line |
 | |
The apoptosis induction potential for vacA&oline/cagA+ strain of H. pylori in AGS cells was 4.43±0.43 for 24 h, 3.71±0.42 for 48 h and 2.52±0.25 for 72 h (Table 1).
The apoptotic activity for vacA&oline/cagA+ strain of H. pylori in HEF cells was 3.83±0.25 for 24 h, 3.49±0.09 for 48 h and 2.05±0.12 for 72 h (Table 2).
Statistically analysis didn’t show a difference between the two cell lines in apoptotic sensitivity (p>0.05). The apoptosis induction potential for cagA&oline/vacA&oline strain of H. pylori in AGS cells was 5.2±0.3 for 24 h, 5.1±0.41 for 48 h and 5.1±0.25 for 72 h. The apoptotic activity for cagA&oline/vacA&oline strain of H. pylori in HEF cells was 5±0.10 for 24 h, 4.9±0.4 for 48 h and 4.9±0.23 for 72 h.
Post hoc test showed that there was no significant difference between cagA&oline/vacA+ and vacA&oline/cagA+ strains in apoptosis induction potential in AGS cells (p>0.05), but a significant difference between cagA&oline/vacA+ and wild type strains of H. pylori (p<0.05) was obtained. In addition, comparison between vacA&oline/cagA+ and wild type to induce apoptosis in AGS showed a distinctive apoptotic (p<0.05). Statically analysis showed that there were significant differences between cagA&oline/vacA+, vacA&oline/cagA+, wild type and double negative strains of H. pylori in apoptosis induction (p<0.05). The results showed that double negative strain of H. pylori could not induce apoptosis in these two cell lines.
PARP cleavage for HEF cells indicate that there was no significant difference between cagA&oline/vacA+ and vacA&oline/cagA+ strains in apoptosis potential, but between cagA&oline/vacA+ and wild type strains (p<0.05) was found. Also, there were significant differences between vacA&oline/cagA+ strain and wild type strain (p<0.05).
General linear model analysis showed that there was a significant difference in apoptosis induction potential by four different strains of H. pylori in various time intervals (24, 48 and 72 h) between AGS and HEF cell lines (p<0.05). In addition, ANOVA showed that all the strains of bacteria tested were varied in apoptosis potential.
Post hoc tests also confirmed that the mean differences of apoptosis potential between the strains were significant especially when the assessment were performed in 48 to 72 h incubation times (p<0.05).
DISCUSSION
The development of intestinal-type gastric adenocarcinoma involves progression through a well-defined series of histological steps, initiated by the transition from normal mucosa to chronic superficial gastritis, followed by the appearance of atrophic gastritis and intestinal metaplasia and, finally, dysplasia and adenocarcinoma (Topal et al., 2004).
Enhanced rates of cell loss during H. pylori infection could potentially accelerate the development of gastric atrophy or intestinal metaplasia (Choi et al., 2003; Covacci et al., 1999; Rokkas et al., 1999).
Gastric adenocarcinoma is strongly associated with the presence of H. pylori (Peek, 2002). One mechanism by which H. pylori may augment the risk for carcinogenesis is by altering cellular turnover. Mucosal hyper proliferation has been demonstrated within H. pylori infected gastric tissue (Rokkas et al., 1999) and multiple studies have concluded that H. pylori is also associated with increased levels of apoptosis in vivo (Jones et al., 1997; Rudi et al., 1998). However, another study has concluded that H. pylori infection is not associated with increased apoptosis (Hirasawa et al., 1999). Explanations for this discordance may include various environmental influences and diversity among infecting H. pylori strains. To investigate the relationship between H. pylori and apoptosis, in vitro assays are very useful because multiple variables can be carefully controlled. This in vitro system provides useful insights into the interaction of H. pylori with gastric epithelial cells in the human stomach. By using an in vitro system to study interactions of H. pylori with mammalian cells, we have shown that different strains of H. pylori induce apoptosis in AGS gastric epithelial cells and HEF fibroblast cells.
The mechanisms by which H. pylori interacts with epithelial cells to induce apoptosis are not known (Shirin, 1998; El-Shahat et al., 2005). There is evidence both for increased expression of the proapoptotic Bcl-2 family member, Bak (Chen et al., 1997) and involvement of the Fas-Fas ligand pathways (Wang et al., 2000). However, other observations suggest the involvement of other pathways in H. pylori induced apoptosis. Further exploration of the molecular mechanisms involved in the effects of H. pylori on cell cycle control may provide insights into the role of this organism in gastric carcinogenesis.
The certain outcome of H. pylori infection in population depends on both genetic characteristics of patients and bacterial strains which could explain the variation in disease process development. One of the mechanisms which need to be clarified is the varied potential of H. pylori strain in destruction of target cells. In the present study we used wild type, either cagA or vacA negative and double negative strains of H. pylori and observed their effects on two different cell lines. Since different apoptotic pathways may operate in different cell types leading to controversial findings, we focused on PARP acting as a joining point for two different hypothetical apoptotic pathways operating in cells. Our in vitro study indicated that direct contact of H. pylori with both gastric cancer cell line and fibroblast cell line was able to induce apoptosis. However, the extent of apoptosis induction was varied either between cell lines or between different bacterial strains.
Based on present results, among different strains of H. pylori, wild type (cagA+/vacA+) strain that had both cagA and vacA genes induced more apoptotic reaction on AGS cell line compared to mutant strains after 72 h (2.1±0.09) and then cagA&oline/vacA+ strain induced apoptosis (2.32±0.61) more than cagA+/vacA&oline strain (2.52±0.25). Double negative strain of H. pylori did not induce apoptosis in compare to time 0 (5.1±0.25). This is in agreement with previous data indicating that induction of apoptosis in gastric epithelial cells either in vivo or in vitro conditions is related to the genotype status of H. pylori strains (Kodoma et al., 1998; Le'Negrate et al., 2001).
In contrast, another study detected apoptosis during infection with both cagA+/vacA+ strains and cagA&oline/vacA&oline H. pylori strains (Wagner et al., 1997).
These discordant findings may due to using different epithelial cells. Wagner and colleagues used different epithelial cells (HM02 cells) for assessing of apoptosis and different epithelial cells may have different patterns in undergoing apoptosis.
In present study we also co-cultured different strains of H. pylori with HEF cell line. Like AGS cells, wild type strains of H. pylori induced apoptosis on HEF cell line more than the others after 72 h (1.46±0.13) and then cagA&oline/vacA+ strain (1.8±0.32) induced apoptosis more than cagA+/vacA&oline strain (2.05±0.12). But, double negative strain of H. pylori did not induce apoptosis in HEF cells after 72 h in compare to time 0 (4.9±0.25). Present finding showed that wild type H. pylori that had both cagA and vacA genes was more potent in induction of apoptosis than the mutant strains.
Table 1 and 2 showed that cagA&oline/vacA+ strains of H. pylori induced apoptosis more than cagA+/vacA&oline strains in both AGS and HEF cell lines. This finding showed that vacA gene is more potent in apoptosis induction than cagA gene.
Present finding is in agreement with Moss et al. (2001) study on cag PAI association with increased apoptosis of gastric epithelial cells Kuck et al. (2001) have shown that VacA is at least one of the H. pylori factors capable of apoptosis induction (Kuck et al., 2001).
Comparison of apoptosis induction in two cell lines by different strains of H. pylori after 72 h showed that HEF cells underwent apoptosis more than AGS cells.
Wagner et al. (1997) showed that cancer cell lines undergo about 50% less apoptosis in response to H. pylori than normal cells.
The present study, showed that apoptosis induction by different strains of H. pylori on HEF cells was more than AGS cells, but, it was not two fold than cancer cell line. Explain for this discrepancy may due to the type of cell line.
Contradictory findings reported by (Maeda et al. (2002) on effect of vacA mutant H. pylori on reduction of cell viability may root from exclusion of other important virulence genes like cagA from the study. However, there are many other controversial reports on potential of cag and vac genes on induction of apoptosis either in vivo or in vitro (Chen et al., 1997; Rudi et al., 1998; Shirin et al., 1999).
In conclusion, the present study showed that different strains of H. pylori but not double negative strain can induce apoptosis, though not up to the same extent on different cell types during a specified time limit. These differences are likely under influence of different bacterial characteristics and also may influence by numerous host derived factors.
REFERENCES
- Anti, M., A. Armuzzi, A. Gasbarrini and G. Gasbarrini, 1998. Importance of changes in epithelial cell turn over during Helicobacter pylori infection in gastric carcinogenesis. Gut, 43: S27-S32.
Direct Link - Ashktorab, H., S. Frank, A.R. Khaled, S.K. Durum, B. Kifle and D.T. Smoot, 2004. Bax translocation and mitochondrial fragmentation induced by Helicobacter pylori. Gut, 53: 805-813.
Direct Link - Chen, G., E.M. Sordillo, W.G. Ramey, J. Reidy and P.R. Holt et al., 1997. Apoptosis in gastric epithelial cells is induced by Helicobacter pylori and accompanied by increased expression of BAK. Biochem. Biophys. Res. Commun 239: 626-632.
Direct Link - Choi, I.J., J.S. Kim and J.M. Kim, 2003. Effect of inhibition extracellular signal-regulated kinase 1 and 2 pathway on apoptosis and bcl-2 expression in Helicobacter pylori infected AGS cells. Infect. Immunol., 71: 830-837.
Direct Link - Conlin, V.S., S.B. Curtis, Y. Zhao, E.D. Moore and V.C. Smith et al., 2004. Helicobacter pylori infection targets adherence junction regulatory proteins and results in increased rates of migration in human gastric epithelial cells. Infect. Immunol., 72: 5181-5192.
Direct Link - Covacci, A., J.L. Telford, G. Del Giudice, J. Parsonnet and R. Rappuoli, 1999. Helicobacter pylori virulence and genetic geography. Science, 284: 1328-1333.
Direct Link - Cover, T.L., U.S. Krishna, D.A. Israel and R.M. Jr. Peek, 2003. Induction of gastric epithelial cell apoptosis by Helicobacter pylori vacuolating cytotoxin. Cancer Res., 63: 951-957.
Direct Link - El-Shahat, M., S. El-Masry and M. Lotfi, 2005. Relationship of Helicobacter pylori to Bcl-2 family expression, DNA content and pathological characteristics of gastric cancer. Int. J. Gastrointestinal Cancer, 36: 61-68.
Direct Link - Guillemin, K., N.R. Salama and L.S. Tompkins, 2002. Cag pathogenicity island-specific responses of gastric epithelial cells to Helicobacter pylori infection. PNAS., 99: 15136-15141.
Direct Link - Hirasawa, R., M. Tatsuta, H. Iishi, H. Yano, M. Baba, N. Uedo and N. Sakai, 1999. Increase in apoptosis and decrease in ornithine decarboxylase activity of the gastric mucosa in patients with atrophic gastritis and gastric ulcer after successful eradication of Helicobacter pylori. Am. J. Gastroenterol., 94: 2398-2402.
Direct Link - Hua, J., P.Y. Zheng and K.G. Yeoh, 2000. The status of the cagA gene does not predict Helicobacter pylori associated peptic ulcer disease. Microbios, 102: 113-120.
Direct Link - Jones, N.L., P.T. Shannon, E. Cutz, H. Yeger and P.M. Sherman, 1997. Increase in proliferation and apoptosis of gastric epithelial cells early in the natural history of Helicobacter pylori infection. Am. J. Pathol., 151: 1695-1703.
Direct Link - Jones, N.L., A.S. Day, H.A. Jennings and P.M. Sherman, 1999. Helicobacter pylori induces gastric epithelial cell apoptosis in association with increased Fas receptor expression. Infect. Immunol., 67: 4237-4242.
Direct Link - Kodoma, K., T. Fujioka and A. Ito, 1998. Toxigenicity of Helicobacter pylori isolates possessing cagA gene and vacuolating cytotoxin. J. Gastroenterol., 33: 14-17.
Direct Link - Kuck, D., B. Kolmerer and C. Iking-Konert, 2001. Vacuolating Cytotoxin of Helicobacter pylori induces apoptosis in the human gastric epithelial cell line AGS. Infect. Immunol., 69: 5080-5087.
Direct Link - Le'Negrate, G., V. Ricci, V. Hofman, B. Mograbi, P. Hofman and B. Rossi, 2001. Epithelial intestinal cell apoptosis induced by Helicobacter pylori depends on expression of the cag pathogenicity island phenotype. Infect. Immunol., 69: 5001-5009.
Direct Link - Maeda, S., H. Yoshida, Y. Mitsuno, Y. Hirata, K. Ogura, Y. Shiratori and M. Omata, 2002. Analysis of apoptotic and antiapoptotic signaling pathways induced by Helicobacter pylori. J. Clin. Pathol., 55: 286-293.
Direct Link - Moss, S.F., E.M. Sordillo, A.M. Abdalla, V. Makarov and Z. Hanzely et al., 2001. Increased gastric epithelial cell apoptosis associated with colonization with cagA+ Helicobacter pylori strains. Cancer Res., 61: 1406-1411.
Direct Link - Peek, R.M. Jr., M.J. Blaser, D.J. Mays, M.H. Forsyth and T.L. Cover et al., 1999. Helicobacter pylori strain-specific genotypes and modulation of the gastric epithelial cell cycle. Cancer Res., 59: 6124-6131.
Direct Link - Peek, R.M. Jr. and M.J. Blaser, 2002. Helicobacter pylori and gastrointestinal tract adenocarcinoma. Nat. Rev. Cancer, 2: 28-37.
Direct Link - Rokkas, T., S. Ladas, C. Liatsos, E. Petridou and G. Papatheodorou et al., 1999. Relationship of Helicobacter pylori CagA status to gastric cell proliferation and apoptosis. Dig. Dis. Sci., 44: 487-493.
Direct Link - Rudi, J., D. Kuck, S. Strand, A. Von Herbay and S.M. Mariani et al., 1998. Involvement of the CD95 (APO-1/Fas) receptor and ligand system in Helicobacter pylori-induced gastric epithelial apoptosis. J. Clin. Invest., 102: 1506-1514.
Direct Link - Shibayama, K., Y. Doi and N. Shibata, 2001. Apoptotic signaling pathway activated by Helicobacter pylori infection and increase of apoptosis-inducing activity under serum-starved conditions. Infect. Immunol., 69: 3181-3189.
Direct Link - Shirin, H., E.M. Sordillo, S.H. Oh, H. Yamamoto, T. Delohery, I.B. Weinstein and S.F. Moss, 1999. Helicobacter pylori inhibits the G1 to S transition in AGS gastric epithelial cells. Cancer Res., 59: 2277-2281.
Direct Link - Topal, D., V. Goral, F. Yilmaz and I.H. Kara, 2004. The relation of Helicobacter pylori with intestinal metaplasia, gastric atrophy and Bcl-2. Turk. J. Gastroenterol., 15: 149-155.
Direct Link - Wagner, S., W. Beil, J. Westermann and R.P.H. Logan, 1997. Regulation of gastric epithelial cell growth by Helicobacter pylori: Evidence for a major role of apoptosis. Gastroenterology, 113: 1836-1847.
Direct Link - Wang, J., X. Fan and C. Lindholm, 2000. Helicobacter pylori modulates lymphoepithelial cell interactions leading to epithelial cell damage through Fas/Fas ligand interactions. Infect. Immunol., 68: 4303-4311.
Direct Link