
Latifeh Pourakbar
Urmia University, Iran
Masoud Khayami
Urmia University, Iran
Jalil Khara
Urmia University, Iran
Tayabeh Farbodnia
Urmia University, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4092-4096







ABSTRACT
Growth parameters and biochemical changes were studied in roots and leaves of 15 day old maize grown in a nutrient solution containing various copper concentrations (0, 25, 50, 75 and 100 μM). An accumulation of H2O2 was observed in roots and shoots. The leaf chlorophyll a, b and carotenoid contents decreased with increasing Cu concentration. The results demonstrated adverse effects of Cu on N metabolism and plant growth. Cu exposure elevated Cu concentration and decreased Nitrate Reductase (NR) activity in the roots and shoots. However, Cu exposure increased total free amino acid content in the leaves.
PDF Abstract XML References Citation
How to cite this article
Latifeh Pourakbar, Masoud Khayami, Jalil Khara and Tayabeh Farbodnia, 2007. Physiological Effects of Copper on Some Biochemical Parameters in Zea mays L. Seedlings. Pakistan Journal of Biological Sciences, 10: 4092-4096.
DOI: 10.3923/pjbs.2007.4092.4096
URL: https://scialert.net/abstract/?doi=pjbs.2007.4092.4096
DOI: 10.3923/pjbs.2007.4092.4096
URL: https://scialert.net/abstract/?doi=pjbs.2007.4092.4096
INTRODUCTION
Plants take up Cu and its accumulation has been reported in leaves, stems, fruits, seeds and roots which increases with increase in the exogenous Cu level (Pinochet et al., 1999; Xiong and Wang, 2005). Although Cu is an essential micronutrient for normal plant metabolism, playing an important role in a large number of metalloenzymes, photosynthesis-related plastocyanin and membrane structure, it has been reported to be among the toxic heavy metals (Wong and Bradshaw, 1982; Li and Xiong, 2004). Excessive Cu accumulated in plant tissue can be toxic to plants, affecting several physiological and biochemical processes and growth (Fernandes and Henriques, 1991). Copper treatment brings change in nitrogen metabolism with a reduction of total nitrogen (Llorens et al., 2000). Cu exposure results in increase of free amino acids (Mazen, 2004) and inhibition of nitrate reductase activity (Luna et al., 1997). The presence of Cu significantly decreases chlorophyll content and inhibits growth (Ralph and Burchett, 1998; Fargasova, 2001).
Most of the in vitro studies using isolated chloroplasts or excised leaves have reported a direct effect of copper on the photosynthetic electron transport chain (Sandman and Böger, 1980; Szalontai et al., 1999). PSII seems to contain binding sites for excess Cu2+ both on the oxidizing and reducing side (Patsikka et al., 2001).
The aim of this study was to investigate the effect of copper toxicity on nitrate reductase activity, H2O2, protein, nitrate, chlorophyll contents and level of hill reaction in Cu-treated maize plant.
MATERIALS AND METHODS
Seeds of Zea mays L. were surface sterilized with 0.1% HgCl2 for 10 min, rinsed thoroughly with running water and germinated in a greenhouse on moist filter paper in Petri dishes for 3 days. The seedlings were randomly placed in polyethylene pots (10 plants per pot) filled with 300 mL of a non-aerated modified Hoagland's nutrient solution (Ouzounidou et al., 1994).
A randomized block, experimental design with five Cu treatments (0, 25, 50, 75 and 100 μM Cu) and three replicates was used. Copper was supplied as CuSO4.5H2O. The plants were grown for 15 days in a growth chamber (16 h light/8 h dark) under a light intensity of 15 W m‾2 at plant level with a day/night temperature regime of 27/22°C and relative humidity of 85% (Ouzounidou et al., 1994). After 15 days, grown seedlings were taken as the reference material for the experiments done in the presence of added copper.
The H2O2 level was colorimetrically measured as described by Jana and Choudhuri (1981).
The soluble protein content was colorimetrically measured as described by the method of Bradford (1976). Protein levels were expressed as mg g‾1 fresh weight.
The chlorophyll a, b and carotenoid content were colorimetrically measured as described by Dere et al. (1998). The amount of these pigments was calculated according to the formulas of Lichtenthaler and Wellburn (1985).
The nitrate content, nitrate reductase (EC 1.6.6.1) activity and total free amino acid content were colorimetrically measured as described by Xiong et al. (2006).
The PSII electron transfer activity (hill reaction) was colorimetrically measured as described by Patsikka et al. (2001). Photoinhibition was monitored by measuring the rate by which the reduce dichlorophenol-indophenol (DCPIP).
All experiments process was conducted in the Urmia university plant physiology laboratory (of September to November 2006).
RESULTS
A significant reduction in shoot and root soluble proteins were observed in Cu-treated plants (Fig. 1).
Figure 2 shows the endogenous H2O2 level roots and shoots of maize seedlings in the presence and absence of CuSO4. It is clear that CuSO4-treated roots in 100 μm concentration of Cu had higher H2O2 level than controls plant and shoots of other cu-treated plant.
Present results demonstrated Cu adverse effects on N metabolism.
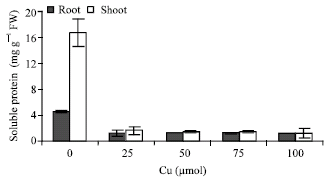 | |
| Fig. 1: | Changes in soluble protein content in shoots and roots of the maize plant in the different Cu concentrations. Vertical bars represent standard errors |
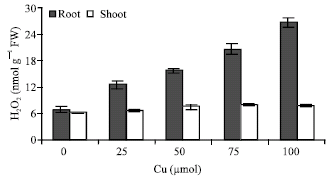 | |
| Fig. 2: | Changes in H2O2 content in shoots and roots of maize plant in the different Cu concentrations. Vertical bars represent standard errors |
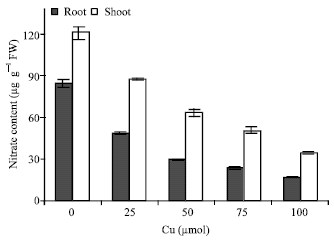 | |
| Fig. 3: | Changes in nitrate content in shoots and roots of maize in the different Cu concentrations. Vertical bars represent standard errors |
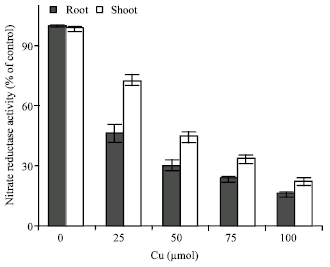 | |
| Fig. 4: | Changes in nitrate reductase activity in shoots and roots of maize in the different Cu concentrations. Vertical bars represent standard errors |
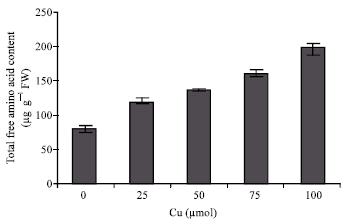 | |
| Fig. 5: | Changes in total free amino acids content in shoots of maize in the different Cu concentrations. Vertical bars represent standard errors |
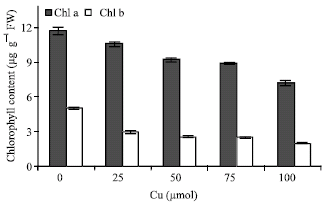 | |
| Fig. 6: | Changes in total chlorophyll (a and b) content in leaves of maize in the different Cu concentrations. Vertical bars represent standard errors |
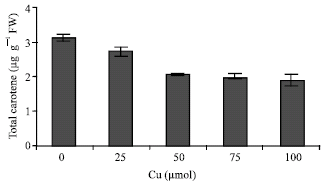 | |
| Fig. 7: | Changes in total carotene content in leaves of maize plant in the different Cu concentrations. Vertical bars represent standard errors |
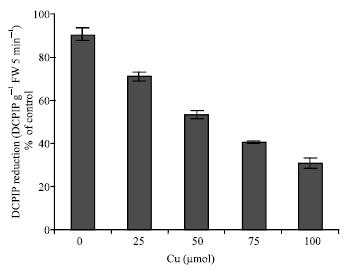 | |
| Fig. 8: | Changes in DCPIP reduction in leaves of maize in the different Cu concentrations. Vertical bars represent standard errors |
Cu exposure decreased nitrate content (Fig. 3) and nitrate reductase activity (Fig. 4) but total free amino acids content in the leaves increased by copper exposure (Fig. 5), indicating a role of free amino acids in detoxification of copper stress.
The shoots were significantly affected by Cu, which resulted in a decline of pigment content. The chlorophyll a and b (Fig. 6) and carotenoid content (Fig. 7) were decreased in Cu-treated plants. Effect of Cu on the DCPIP reduction shows in Fig. 8. DCPIP reduction decreased with increasing Cu concentrations. It is clear that DCPIP reduction in 100 μM Cu-treated plants had about 70% lower than controls plant.
DISCUSSION
Cu2+ is required by biological systems as a structural and catalytic enzyme component and in the soil Cu2+ can be a stress factor by causing physiological responses that can decrease the vigour of the plants and inhibit plant growth (Ouzounidou, 1994).
The soluble protein content in plant cells is an important indicator of their physiological state. The reduced amount of soluble protein in the organs of the Cu-treated maize plants was most probably a result of the reduced biosynthesis or the accelerated catabolic processes, as well. This suggestion is also confirmed by Vassilev (1997) in his study on Cd and Stoeva and Bineva (2003) in their study on As (Arsenic), according to whom, the protein degradation to amino acids is in fact an adaptation of the cells to the carbohydrate deficiency. On the other hand, the accelerated catabolism is probably due to the considerable disturbances in the membrane systems, in response to the metal phytotoxicity. Present results demonstrated Cu adverse effects on N metabolism. Cu exposure decreased nitrate reductase activity and total chlorophyll content. Total free amino acids content in the leaves increased by copper exposure, indicating a role of free amino acids in detoxification of copper stress. The results suggested that Cu toxicity to the plant was at least partly via influence on N metabolism. Reduction of nitrate to nitrite catalyzed by Nitrate Reductase (NR) is considered the rate-limiting step in the assimilation of nitrate (Campbell, 1999), which eventually affects the growth and organic nitrogen status of the plants. Another direct effect of Cu exposure on NR activity decrease may lie in an increase in the enzyme breakdown induced by toxic oxygen species generated during Cu stress. Free radicals can cause the breakdown of proteins by oxidative reaction or by increasing proteolytic activity (Salo et al., 1990). It suggests that Cu-mediated generation of reactive oxygen species (e.g., H2O2, O2• or OH•) could have occurred. These radicals could attack nitrate reductase and cause a decrease of its activity. In addition, NR activity decrease under Cu exposure may involve the damage of Cu to the plants photosynthesis. In present experiments, the chlorophyll content was significantly reduced by Cu treatment. This suggests that the plants photosynthesis could have been damaged because of the adverse effects of Cu on chlorophyll and thus supports the possibility of an indirect effect of photosynthesis damage on NR activity by Cu exposure. Our experiments indicated that NR activity in leaves was much higher than that in the roots. Two possibilities may be considered to explain these observations. The first may lie in a concentration effect of Cu the higher the Cu concentration in the roots the more serious would be the inhibition of the enzyme. The second may suggest that the reduction process from nitrate to nitrite catalyzed by NR has occurred mainly in leaves (Xiong et al., 2006).
The photosynthetic pigments are some of the most important internal factors, which in certain cases are able to limit the photosynthesis rate. It was established that total carotene (Car) decreased to a lesser extent than Chl. The decreased Chl and Car may be due to the fact that there is an oxidative stress, which is a marker of the tissue aging, as result of the stress factors of the environment (Hendry and Grime, 1993). A reduction of the pigment content in the case of increasing levels of heavy metals and metalloids, was also established by Bogoeva (1998). In the present study, Cu treatment also was found to reduce total chlorophyll content. This may result from chlorophyll degradation and chlorophyll synthesis inhibition due to Cu-induced inhibition of aminolaevulinic acid dehydratase (ALA-dehydrates) (Fernandes and Henriques, 1991). Also, chlorophyll content decline could result from an indirect adverse effect of Cu exposure on N assimilation of the plants, since biosynthesis of chlorophylls is dependant on N supply from the plants’ N assimilation. In this respect, Cu exposure could cause decrease of N assimilation, thus leading to a decline in chlorophyll content.
Free-radical damage induced by heavy metals by maintaining a more reducing environment in plant cells (Surasak et al., 2002). Hydrogen peroxide (H2O2) is a constituent of oxidative plant metabolism. It is a product of peroxisomal and chloroplastic oxidative reactions (Del Rio et al., 1992). H2O2 itself is an active oxygen species. H2O2 can also react with superoxide radicals to form more reactive hydroxyl radicals in the presence of trace amounts of Fe or Cu (Thompson et al., 1987). The hydroxyl radicals initiate self-propagating reactions leading to peroxidation of membrane lipids and destruction of proteins (Asada and Takahashi, 1987; Bowler et al., 1992; Halliwell, 1987).
Copper is essential for electron transport not only by being part of plastocyanin, but probably also as a stabilizer of the lipid environment close to electron carrier of PSII complex (Patsikka et al., 2001). However, excess Cu2+ has also harmful effects on the donor and acceptor side of PSII (Yruela et al., 1996). Cu2+-induce loss of PSII activity is enhanced by light which suggests involvement of photoinhibition (Patsikka et al., 1998). The ability of an exogenous electron donor to partially restore PSII activity lost by illumination of thylakoids in the presence of Cu2+ suggests that the damage started at the donor side of PSII (Patsikka et al., 2001). In the absence of electron donation from oxygen-evolving complex, the long lived P680* oxidizes nearby components of PSII (Bumann and Oesterhelt, 1995). Furthermore Jegerschod et al. (1995) have suggested that the binding of the Cu2+ near YZ would slow down the reduction of the oxidized primary electron donor, P680*. When the plants are grown under copper additions, the inhibitory effect of Cu2+ on the photosynthetic machinery is cumulative in the course of time and the extent of inhibition may also depend on the physiological stage of the leaf studied (Maksymiec and Baszynsky, 1999).
REFERENCES
- Bogoeva, I., 1998. Pigment content in cereal crops growth on copper polluted soil. Bulgar J. Agric. Sci., 6: 767-772.
Direct Link - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Bumann, D. and D. Oesterhelt, 1995. Destruction of a signal cholorophyll is correlated with the photoinhibition of photosystem II whit a transiently inactive donor side. Proc. Nat. Acad Sci. USA., 92: 12195-12199.
Direct Link - Campbell, W.H., 1999. Nitrate reductase structure, function and regulation: Bridging the gap between biochemistry and physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 277-303.
Direct Link - Del-Rio, L.A., L.M. Saudalio, J.M. Palma, P. Bueno and F.J. Corpas, 1992. Metabolism of oxygen radicals in peroxisomes and cellular implications. Free Radical Med. Biol., 13: 557-580.
Direct Link - Dere, A., T. Gunes and R. Sivaci, 1998. Spectrophotometric determination of Chlorophyll-A, B and total carotenoid contents of some algae species using different solvents. Truk. J. Bot., 22: 13-17.
Direct Link - Fargasova, A., 2001. Phytotoxic effects of Cd, Zn, Pb, Cu and Fe on Sinapis alba L. seedling and their accumulation in roots and shoots. Biol. Plant, 44: 471-473.
Direct Link - Fernandes, J.C. and F.S. Henriques, 1991. Biochemical, physiological and structural effects of excess copper in plants. Bot. Rev., 57: 246-273.
CrossRefDirect Link - Hendry, G.A.F. and J.P. Grime, 1993. Methods in Comparative Plant Ecology: A Laboratory Manual. Chapman and Hall, UK pages: 252.
Direct Link - Jena, S. and M.A. Choudhuri, 1982. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat. Bot., 12: 345-354.
CrossRefDirect Link - Jegerschod, C., J.B. Arellano, W.P. Schroder, P.J.M. Vankan, M. Barton and S. Styring, 1995. Copper (II) inhibition of electron transfer through photosystem II studied by EPR spectroscopy. Biochemistry, 34: 12747-12754.
Direct Link - Li, T. and Z.T. Xiong, 2004. A novel response of wild-type duckweed (Lemna paucicostata Hegelm.) to heavy metals. Environ. Toxicol., 19: 95-102.
Direct Link - Lichtenthaler, H.K. and A.R. Wellburn, 1985. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biol. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Llorens, N., L. Arola, C. Blade and A. Mas, 2000. Effects of copper exposure upon nitrogen metabolism in tissue cultured Vitis vinifera. Plant Sci., 160: 159-163.
CrossRefPubMedDirect Link - Luna, C.M., L.M. Casano and V.S. Trippi, 1997. Nitrate reductase is inhibited in leaves of Triticum aestivum treated with high levels of copper. Physiol. Plant., 101: 103-108.
CrossRefDirect Link - Mazen, A.M.A., 2004. Accumulation of four metals in tissues of Corchorus olitorius and possible mechanisms of their tolerance. Biol. Plant, 48: 267-272.
Direct Link - Ouzounidou, G., 1994. Copper-induced changes on growth, metal content and photosynthetic function of Alyssum montanum L. plants. Environ. Exp. Bot., 34: 165-172.
CrossRefDirect Link - Ouzounidou, G., M. Ciamporova, M. Moustakas and S. Karataglis, 1995. Responses of maize (Zea mays L.) plants to copper stress-I growth, mineral content and structure of roots. Environ. Exp. Bot., 35: 167-176.
Direct Link - Patsikka, E., E.M. Aro and E. Tyystjarvi, 1998. Increase in the quantum yield of photoinhibition contributes to copper toxicity in vivo. Plant Physiol., 117: 619-627.
PubMedDirect Link - Patsikka, E., E.M. Aro and E. Tyystjarvi, 2001. Mechanism of copper-enhanced photoinhibition in thylakoid membranes. Physiol. Plant., 113: 142-150.
Direct Link - Pinochet, H., I. De Gregori, M.G. Lobos and E. Fuentes, 1999. Selenium and copper in vegetables and fruits grown on long-term impacted soils from Valparaiso Region, Chile. Bull. Environ. Contam. Toxicol., 63: 327-334.
Direct Link - Ralph, P.J. and M.D. Burchett, 1998. Photosynthetic response of Halophila ovalis to heavy metal stress. Bull. Environ. Contam. Toxicol., 113: 91-101.
Direct Link - Salo, D.C., R.E. Pacifici, S.W. Lin, C. Giulivi and K.J.A. Davies, 1990. Superoxide dismutase undergoes proteolysis and fragmentation following oxidative modification and inactivation. J. Biol. Chem., 265: 11919-11927.
Direct Link - Stoeva, N. and T. Bineva, 2003. Oxidative changes and photosynthesis in OAT plants growth in AS-contaminated soil. Bulg. J. Plant Physiol., 29: 87-95.
Direct Link - Siripornadulsil, S., S. Traina, D.P.S. Verma and R.T. Sayre, 2002. Molecular mechanisms of proline-mediated tolerance to toxic heavy metals in transgenic microalgae. Plant Cell., 14: 2837-2847.
Direct Link - Szalontai, B., L.I. Horvath, M. Debreczeny, M. Droppa and G. Horvath, 1999. Molecular rearrangements of thylakoids after heavy metal poisoning, as seen by Fourier Transform Infrared (FTIR) and Electron Spin Resonance (ESR) spectroscopy. Photosynth. Res., 61: 241-252.
Direct Link - Xiong, Z.T. and H. Wang, 2005. Copper toxicity and bioaccumulation in Chinese cabbage (Brassica pekinensis Rupr.). Environ. Toxicol., 20: 188-194.
Direct Link - Xiong, Z.T., L. Chao and G. Bing, 2006. Phytotoxic effects of copper on nitrogen metabolism and plant growth in Brassica pekinensis Rupr. Ecotoxicol. Environ. Saf., 64: 273-280.
Direct Link - Yruela, I., J.J. Pueyo, P.J. Alonso and R. Picorel, 1996. Photoinhibition of photosystem II from higher plants. Effect of copper inhibition. J. Biol. Chem., 271: 27408-27415.
Direct Link