
H. Pakniyat
Department of Crop Production and Plant Breeding, College of Agriculture, Shiraz University, Shiraz, Iran
M. Armion
Department of Crop Production and Plant Breeding, College of Agriculture, Shiraz University, Shiraz, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4081-4086







ABSTRACT
Twenty eight sugar beet genotypes were analysed for their tolerance at 3 NaCl levels (0, 3000 and 6000 mg NaCl kg‾ 1 soil) and Na+, K+, Na+/K+ and free proline were measured from the leaf samples. Results showed that increasing salinity level caused an increase in Na+, Na+/K+ and proline, but a decrease in K+ content of leaf samples (p ≤0.01). As compared to non-tolerant genotypes, tolerant ones accumulated more Na+ and Na+/K+ and proline and less K+. It seems that Na+ and proline accumulation in shoots are effective mechanisms for osmotic pressure adjustment and plant tolerance to salinity, a mechanism commonly seen in sugar beet ancestors.
PDF Abstract XML References Citation
How to cite this article
H. Pakniyat and M. Armion, 2007. Sodium and Proline Accumulation as Osmoregulators in Tolerance of Sugar Beet Genotypes to Salinity. Pakistan Journal of Biological Sciences, 10: 4081-4086.
DOI: 10.3923/pjbs.2007.4081.4086
URL: https://scialert.net/abstract/?doi=pjbs.2007.4081.4086
DOI: 10.3923/pjbs.2007.4081.4086
URL: https://scialert.net/abstract/?doi=pjbs.2007.4081.4086
INTRODUCTION
Salinity is a serious environmental constraint to crop production in many parts of the world and the development of crops with improved salt tolerance is proposed as part of solution to this problem (Zhu, 2001). Plants follow different behaviors to combat salinity. Detailed reviews about salt tolerance mechanisms in different species are presented by Ashraf (2004) and Sairam and Tyagi (2004).
Osmotic adjustment has undoubtedly gained considerable recognition as a significant and effective mechanism of salinity tolerance in crop plants. Osmoregulatory effects of proline, glycine betaine and ions on water balance and salt tolerance, have been shown in spinach (Martino et al., 2003), wheat (Abdel-Aziz and Reda, 2000), bean (Shabala et al., 2000), cowpea (Freitas et al., 2001), sugar beet (Ghoulam et al., 2002; Heuer et al., 1981) and a halophyte sea aster (Ueda et al., 2003).
Sugar beet (Beta vulgaris L., family; Chenopodiaceae), has halophytes as ancestors. Its tolerance threshold to salinity is high (7 dS m‾1) (Katerji et al., 1997). Low tolerance to salinity is observed during seed germination and seedling emergence, but there are variations between sugar beet genotypes at high salt stresses (Sadeghian et al., 2000; Ghoulam et al., 2002).
Members of Chenopodiaceae including sugar beet can combat salinity by having osmotic regulating mechanisms due to accumulation of Na+ and Cl‾ in their vacuoles and cytoplasm (Subbarao et al., 2001; Ghoulam et al., 2002). Sugar beet genotypes absorb Na+ and accumulate it in their leaf tissue for regulation and adaptation of its osmotic potential with soil (Flowers, 1988). This may be the reason for considering sugar beet as a tolerant crop.
Although considerable research has been devoted to quantify the salt tolerance of the crop, data are usually based on comparisons among only a few cultivars and survey on a wide range of genotypes and wild progenitors of cultivated species have not been examined or exploited at all. The purpose of this experiment is to study the effect of Na+, K+, Na+/K+ and proline accumulation in 28 sugar beet genotypes, comparison of their salt tolerance and determination of osmotic adjustment role of Na+, Na+/K+ and proline in tolerant and non-tolerant genotypes.
MATERIALS AND METHODS
Twenty eight sugar beet genotypes (Table 1) were compared at 3 salinity levels (0, 3000 and 6000 mg NaCl kg‾ 1 soil) for their Na+, K+, Na+/K+ and proline contents in a completely randomized design with 3 replications. The experiment was conducted in a glasshouse at Agricultural College, Shiraz University in Badjgah, Iran in 2002. These genotypes were random regarding their salinity tolerance and some are planted by local formers in different part of Iran. Genotypes No. 2, 7 and 12 are known as salt tolerant and genotypes No. 4, 6 and 24 are known as non-tolerant ones. These genotypes were used as check genotypes in this experiment.
| Table 1: | Sugar beet genotypes used for their salt tolerance determination in the experiment |
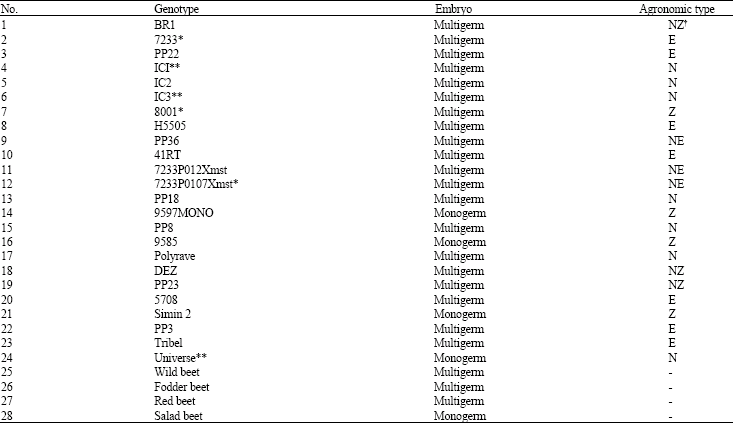 | |
| *: Tolerant check genotypes, **: Non-tolerant check genotypes, χ : N = Normal, Z = Zucker, E = Ernte | |
Ten germinated seed were planted in 5 kg pots filled with a sandy clay loam soil (25% clay, 24% silt and 51% sand, EC = 0.57 dS m‾ 1).
Salt stress treatments were applied 4 weeks after planting and at 4 leaves stage. Salt (NaCl) stock solution was added to appropriate pots in 3 stages to final concentration based on soil field capacity. Four weeks after applying salt treatments (EC = 10 and 15 dS m‾1 for 3000 and 6000 mg NaCl kg‾1 soil, respectively), leaf samples were collected for Na+, K+ and proline measurements.
Na+ and K+ measurements: Leaves were cut from soil surface and, dried at 65°C oven for 48 h and made into fine powder using mortar. Samples (0.5 g) were ashed by putting them into crucibles and placed in 500°C electric furnace, 5 mL of 2N HCl were added to ash samples and mixed with boiling distilled water and filtered by filter paper (Whatman No. 2) into 50 mL volumetric flasks. Na+ and K+ were measured using flame photometer and reported as mg g‾1 dry weight.
Proline measurement: 0.5 g of leaf powder were mixed thoroughly in 10 mL sulfosalicilic acid (3%) and filtered by filter paper (Whatman No. 2) and the proline content was measured by Bates method (Bates et al., 1973).
Statistical analysis: Data were analysed using MSTATC and SPSS softwares. Correlation coefficients for the measured characters were also determined (Table 3). Graphical representations and diagrams were made from EXCEL software.
RESULTS AND DISCUSSION
Na+ content: Increasing salinity levels, increased leaf Na+ contents of the genotypes (Fig. 1a). But the genotypes differed significantly (p<0.01) and the mean Na+ content for tolerant genotypes (2, 7 and 12) were higher than non-tolerant (4, 6 and 24) ones (Fig. 2b and Table 2). Genotypes No. 23, 21, 19, 16, 20 and 9 were not significantly different regarding their Na+ content and they may be considered as salt tolerant genotypes. On the other hand, genotypes No. 25, 1, 18, 26, 3, 15, 28 and 13 which have Na+ contents within the range of non-tolerant check genotypes, may be considered as non-tolerant ones (Table 2). This variation in mechanism of Na+ uptake could be due to some multiple adaptation to toxic ions operating concurrently within a specific plant (Tester and Davenport, 2003; Carden et al., 2003).
This is typical response of Chenopodiaceae family (halophytes), in which plant regulates its tissues osmotic potential by Na+ accumulation and combating with salinity (Eisa and Ali, 2001).
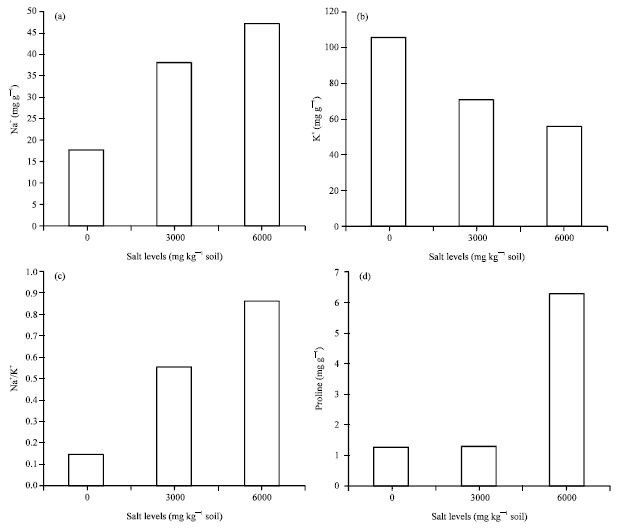 | |
| Fig. 1: | Mean shoot Na+, K+, Na+/K+ and proline content of 28 sugar beet genotypes at 3 salt levels (0, 3000 and 6000 mg kg‾ 1 soil) |
| Table 2: | Shoot Na+, K+, Na+/K+ and proline contents of 28 sugar beet genotypes under salinity (6000 mg NaCl kg‾ 1 soil) ranked on the basis of their Na+ contents |
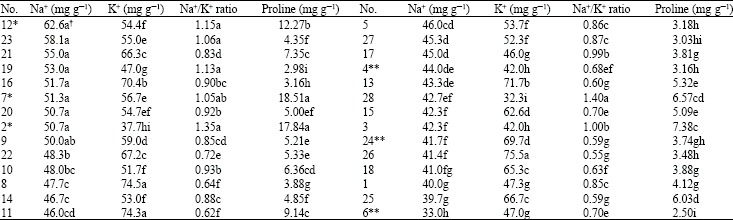 | |
| *: Tolerant check genotypes, **: Non-tolerant check genotypes, χ : Means followed by the same letter(s) (a-i) in each column are not significantly different (DMRT, p<0.01) | |
This reaction is opposite to barley (a glycophyte crop) which is Na+ excludant and in which salt tolerant genotypes accumulate less Na+ in their shoots (Pakniyat et al., 1997, 2003). It seems that in tolerant sugar beet genotypes simultaneous expression of proteinous vector of tonoplast membrane (H+-ATPase port and Na+/H+ antiport) in cell vacoule of the leaves of tolerant genotypes are more than non-tolerant ones (Kirsch et al., 1996; Shi et al., 2002; Parks et al., 2002).
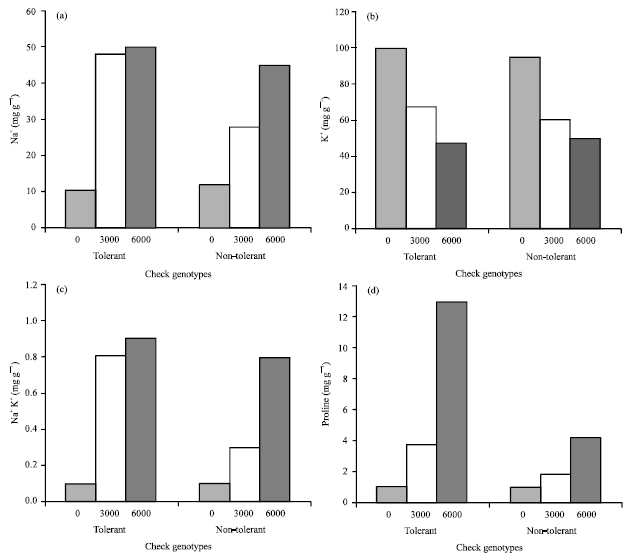 | |
| Fig. 2: | Variation in mean Na+, K+, Na+/K+ and proline contents between tolerant (2, 7 and 12) and non-tolerant (4, 6 and 24) check sugar beet genotypes at different salinity levels (0, 3000 and 6000 mg kg‾ 1 soil) |
| Table 3: | Correlation coefficients between Na+, K+, Na+/K+ and proline contents of sugar beet genotypes under salinity stress |
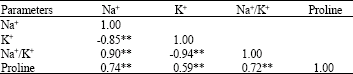 | |
| **: Significant at 1% probability level | |
K+ content: Results showed that by increasing salinity level, mean K+ content of the leaves of the genotypes were decreased and there were significant differences between salinity levels (p<0.01) in this regard (Fig. 1b). K+ content was least for the third level of salinity (6000 mg kg‾ 1) and mean K+ content for non-tolerant genotypes (2, 4 and 6) were higher than tolerant ones (2, 7 and 12) (Fig. 2b and Table 2). These trends have been observed in previous studies also (Flowers, 1985; Warne et al., 1990). This may be due to Na+ role in regulation of sugar beet leaf osmotic potential (Lindhauer et al., 1990) and substitution of Na+ with K+ in this regard.
Negative correlation coefficient between leaf Na+ and K+ content was high and highly significant (R = -0.85, p<0.01) (Table 3). This agrees with the findings of Eisa and Ali (2001) results, which showed a negative linear correlation between these two ions after salt stress in sugar beet leaves. Increasing Na+ accumulation and reduction of K+ content shows a critical role of Na+ in osmotic potential adjustment of sugar beet under salt stress.
Leaf Na+/K+: Results indicated a significant Na+/K+ ratio in leaves (p<0.01) of the genotypes with increasing salinity levels (Fig. 1c). Significant differences (p<0.01) were observed between sugar beet genotypes for their leaves Na+/K+ contents (Table 2). This ratio was higher in tolerant genotype (2, 7 and 12) compared to non-tolerant ones (4, 6 and 24) (Fig. 2c). Genotypes 2, 12 and 28 had the highest leaf Na+/K+ ratios, of which genotypes No. 2 and 12 were among the check, known tolerant genotypes to salinity.
Proline content: Data on proline measurements indicated that proline accumulation has been the consequence of salinity and different salinity levels had significant effects (p<0.01) on leaf proline content of sugar beet genotypes. It seems that plant response to proline drastically increases at 6000 mg NaCl/kg soil (Fig. 1d). Genotypes were significantly different (p<0.01) regarding their leaf proline content under salinity (Table 2). Comparing proline content of tolerant and non-tolerant genotypes, it was indicated that tolerant genotypes accumulated higher level of proline than non-tolerant ones (Fig. 2d). This is in conformity with Gzick (1996) who concluded that higher proline level under salt stress is related to osmotic potential regulation in sugar beet.
In general salt tolerant sugar beet genotypes combat Na+ toxicity by its accumulation in leaf cell vacuoles and regulate their osmotic potential under salinity stress. Besides, these genotypes accumulate higher level of proline in their leaf for their osmotic potential regulation. These findings are in agreement with those investigated in Atriplex which is a halophyte and belongs to Chenopodiaceae family (Glenn et al., 1994).
Correlation coefficients: Correlation coefficients between measured characters are shown in Table 3. There was positive and significant correlations between sugar beet shoot Na+ and proline content (r = 0.74, p<0.01) (Table 3). These 2 measurements can be used for screening tolerant genotypes under salt stress.
Correlation between Na+ and K+ was negatively significant (r = -0.85, p<0.01) which shows osmoregulation of Na+ ion under saline condition in tolerant genotypes and its substitution with K+ (Table 3).
CONCLUSIONS
The most tolerant genotype in this experiment was No. 12, which is among the tolerant check genotypes and the most non-tolerant genotypes was No. 6 which is among the non-tolerant check genotype. These two genotypes had the most and the least amount of Na+ contents.
Regarding proline content, these genotypes were among the group genotypes which contained the highest and the lowest amount of proline content, respectively. Other tolerant and non-tolerant check genotypes followed the same trend regarding their Na+ and proline content. These evidences confirm the osmoregulation role of Na+ and proline in osmotic potential adjustment of the plant and hence its salt tolerance.
We may have crosses between genotypes 12 and 6 to obtain a segregating population to study quantitative trait loci involved in salt tolerance in the future.
REFERENCES
- Ashraf, M., 2004. Some important physiological selection criteria for salt tolerance in plants. Flora-Morphol. Distrib. Funct. Ecol. Plants, 199: 361-376.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Carden, D.E., D.J. Walker, T.J. Flowers and A.J. Miller, 2003. Single cell measurement of the contributions of cytosoloic Na+ and K+ to salt tolerance. Plant Physiol., 131: 676-683.
Direct Link - Ghoulam, C., A. Foursy and K. Fares, 2002. Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot., 47: 39-50.
Direct Link - Gzik, A., 1996. Accumulation of proline and pattern of α-amino acids in sugar beet plants in response to osmotic, water and salt stress. Environ. Exp. Bot., 36: 29-38.
CrossRefDirect Link - Heuer, B. and Z. Plaut, 1989. Photosynthesis and osmotic adjustment of two sugar beet cultivar grown under saline conditions. J. Exp. Bot., 40: 437-440.
CrossRefDirect Link - Katerji, N., J.W. Van-Hoorn, A. Hamdy and M. Mastroili, 1997. Osmotic adjustment of sugar beet in response to soil salinity and its influence on stomatal conductance, growth and yield. Agric. Water Manage., 34: 557-569.
Direct Link - Kirsch, M., A. Zhigang, R. Viereck, R. Low and T. Rausch, 1996. Salt stress induces an increased expression of V-type H+-ATPase in mature sugar beet leaves. Plant Mol. Biol., 32: 543-547.
Direct Link - Lindhauer, M.G., H.G. Header and H. Beringer, 1990. Osmotic potential and solute concentration in sugar beet plants cultivated with varying potassium/sodium ratios. Z. Pflanzenernahr. Bodenk, 153: 25-32.
Direct Link - Di-Martino, C., S. Delfine, R. Pizzuto, F. Loreto and A. Fuggi, 2003. Free amino acids and glycine betaine in leaf osmoregulation of spinach responding to increasing salt stress. New Phytol., 158: 455-463.
CrossRefDirect Link - Pakniyat, H., L.L. Handley, W.T.B. Thomas, T. Connolly, M. Macaulay, P.D.S. Caligari and B.P. Forster, 1997. Comparison of shoot dry weight, Na+ content and C13 values of ari-e and other semi-dwarf barley mutants under salt stress. Euphytica, 94: 7-14.
CrossRefDirect Link - Parks, G.E., M.A. Dietrich and K.S. Schumaker, 2002. Increase vacuolar Na+/H+ exchange activity in Salicornia bigelovii Torr. in response to NaCl. J. Exp. Bot., 53: 1055-1065.
Direct Link - Sadeghian, S.Y., H. Fazli, R. Mohammadian, D.F. Taleghani and M. Mesbah, 2000. Genetic variation for drought stress in sugar beet. J. Sugar Res., 37: 35-77.
Direct Link - Sairam, R.K. and A. Tyagi, 2004. Physiology and molecular biology of salinity stress tolerance in plants. Curr. Sci., 86: 407-421.
Direct Link - Shabala, S., O. Babourina and I. Newman, 2000. Ion-specific mechanisms of osmoregulation in bean mesophyll cells. J. Exp. Bot., 51: 1243-1253.
Direct Link - Shi, H., F.J. Quintero, J.M. Pardo and J.K. Zhu, 2002. The putafive plasma membrance Na+/H+ antiporter SOSI controls long-distance Na+ transport in plants. Plant Cell, 14: 465-477.
Direct Link - Subbarao, G.V., R.M. Wheeler, L.H. Levine and G.W. Stutte, 2001. Glycine betaine accumulation, ionic and water relations of red beet at contrasting levels of sodium supply. J. Plant Physiol., 158: 767-776.
Direct Link - Tester, M. and R. Davenport, 2003. Na+ tolerance and N+ transport in higher plants. Ann. Bot., 91: 503-527.
Direct Link - Ueda, A., M. Kanechi, Y. Uno and N. Inagaki, 2003. Photosynthetic limitations of a halophyte sea aster (Aster tripolium L.) under water stress and NaCl stress. J. Plant Res., 116: 65-70.
PubMed - Warne, P., R.D. Guy, L. Roltins and D.M. Reid, 1990. The effect of sodium sulphate and sodium chloride on growth, morphology, photosynthesis and water use efficiency of Chenopodium rubrum. Can. J. Bot., 68: 999-1006.
Direct Link