
A. Ahmadi
Department of Agronomy and Plant Breeding,
University College of Agriculture and Natural Resources, Tehran University, Karaj, Iran
A. Sio-Se Mardeh
College of Agriculture, Kurdistan University, Sanandaj, Iran
K. Poustini
Department of Agronomy and Plant Breeding,
University College of Agriculture and Natural Resources, Tehran University, Karaj, Iran
M. Esmailpour Jahromi
Department of Agronomy and Plant Breeding,
University College of Agriculture and Natural Resources, Tehran University, Karaj, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4043-4049







ABSTRACT
Insufficient seedling stand establishment is one of the major obstacles to achievement of winter wheat (Triticum aestivum) potential yield. In the present study, seeds of eleven wheat cultivars differing in drought resistance were subjected to hydropriming with distilled water and osmopriming with PEG 6000 for 12 h. Speed of emergence, vigor index and seedling dry weight were studied under two temperature conditions at 11 and 18° C in plant growth incubators. Water stress started 15 Days After Sowing (DAS). As compared with osmopriming, hydropriming clearly improved speed of emergence, vigor index and seedling dry weight. The results were more evident at 11° C indicating that hydropriming is more efficient for cold rather than temperate area. At both temperatures, PEG treatment severely diminished above mentioned traits. At 11° C, drought resistant cultivars i.e., Sardari, Agosta-Sefid, Azar 2 and Sabalan had higher seedlings dry weight as compared with susceptible ones. Therefore, it is proposed that measurement of seedling dry weight may be a proper approach for early screening of wheat drought resistant genotypes.
PDF Abstract XML References Citation
How to cite this article
A. Ahmadi, A. Sio-Se Mardeh, K. Poustini and M. Esmailpour Jahromi, 2007. Influence of Osmo and Hydropriming on Seed Germination and Seedling Growth in Wheat (Triticum aestivum L.) Cultivars under Different Moisture and Temperature Conditions. Pakistan Journal of Biological Sciences, 10: 4043-4049.
DOI: 10.3923/pjbs.2007.4043.4049
URL: https://scialert.net/abstract/?doi=pjbs.2007.4043.4049
DOI: 10.3923/pjbs.2007.4043.4049
URL: https://scialert.net/abstract/?doi=pjbs.2007.4043.4049
INTRODUCTION
Food shortage is one of the most serious global problems in this century and hence to meet the expanding food demands of the rapid growing world population, crop grain production will need to increase (Pimentel and Pimentel, 2006). Considering its highest cultivation area and amount of production amongst crops and being main source of calories for people, wheat (Triticum aestivum) is the most important and strategic cereals in most of developing countries.
Insufficient stand establishment of winter wheat is one of the major problems in the low-precipitation (<300 mm annual) dryland region (Giri and Schillinger, 2003). Wheat seeds cultivated in this area often are exposed to adverse environmental conditions such as drought and cold stresses during germination and seedling emergence. In recent decades interest has grown in the use of priming techniques to ensure rapid and uniform seedling emergence and consequently, an adequate stand establishment of vigorous plants which in turn can affect the yield and quality of final product. Priming has been described as a preplant physiological seed conditioning that can improve seed performance by reducing the time to germination and seedling emergence and also by increasing the tolerance of seeds to stress conditions, such as lack of water and adverse temperatures (Caseiro et al., 2004).
Literature well documents that physiological and biochemical changes including early DNA replication, increased RNA and protein synthesis, increased α-amylase activity, greater ATP availability and energy of germination, faster embryo growth, repair of deteriorated seed parts and reduced leakage of metabolites take place during the seed priming which allow seeds to begin the germination sequences before sowing and consequently, improve emergence (Ruan et al., 2002; Giri and Schillinger, 2003; Basra et al., 2005; Farooq et al., 2006).
Among seed priming techniques, two broad classes may be employed: hydropriming that uses only water in the process of controlled imbibitions and Osmopriming, which involves osmotic solutions to hydrate the seeds (Caseiro et al., 2004). According to Hussain et al. (2006) hydropriming and osmopriming with NaCl are the most promising priming techniques so that they decreased mean emergence time and increased final emergence and achene yield of hybrid sunflower. However, the way in which the priming treatment is done may substantially influence the results. For instance, while seed priming with PolyEthylene Glycol (PEG) had adverse effect on shoot and root length of sunflower (Kayaa et al., 2006), it was the most beneficial treatment to improve asparagus seeds performance (Bittencourt et al., 2005). Generally, the major effect of seed priming on seedling growth resulted from earlier germination which gave the seedlings more time to grow. It has been reported that wheat seed hydropriming for less than 12 h is more effective than other priming methods (Giri and Schillinger, 2003) so that it can increase the rate and extent of emergence, improve seedling vigor, advance flowering and maturity and increase yield (Harris, 2006). Seed priming with potassium salts (1%) for 18 h increased wheat yield under dryland condition (Paul and Choudhury, 1991). In India, seed hydropriming of rice (Oryza sativa L.) improved seedling emergence, green status and drought resistance in a field (Harris et al., 1999). Ruan et al. (2002) reported that rice seed priming treatments increased germination percentage, emergence rate, seedling vigor and establishment and reduced Mean Germination Time (MGT) as compared with non primed seeds. Osmotic solutions enable primed seeds to germinate in a lower temperature than non primed seeds do. According to Bodsworth and Bewley (1981) PEG solution with-10 bar osmotic potential at 10° C for two days had the most positive effect on wheat seed germination at low temperatures. Rice primed seeds with PEG had significantly higher seedling vigor and cold resistance as compared with non primed seeds (Qin and Zheng, 1994). Giri and Schillinger (2003) found that priming treatments enhanced emergence of the wheat cultivars with moderate emergence capability but not the one with strong emergence characteristics; therefore the effect of priming treatments on wheat emergence may be cultivar dependent.
The present study was therefore, carried out with the objectives to evaluate the effect of two priming treatments and temperatures on seed germination and seedling growth and dry weight of wheat cultivars differing in drought resistance under water stress and non-stress conditions.
MATERIALS AND METHODS
Experimental site: The study was conducted at seed control and registration lab of Gharizah research center, Sanandaj, Iran in 2002. The experiment was laid out factorial arranged in a Randomized Complete Block Design (RCBD) with three replications.
Seed materials: Eleven wheat cultivars differing in drought resistance (Sio-Se Mardeh et al., 2006) including Alvand, Cross-Shahi, Mahdavi, M-75-7, Toos, Roushan-Back cross, Glinson (drought susceptible cultivars), Sardari, Sabalan, Agosta-Sefid and Azar 2 (drought resistant ones) were employed. The initial seed moisture content was 12.7% on dry weight basis.
Priming treatments:
| • | Hydropriming: Seeds of each cultivar were placed in Petri dishes containing two layers of filter paper and distilled water at 10° C for 12 h. |
| • | Osmopriming. Seeds of each cultivar were primed in -10 bar PEG 6000 calculated using the equation developed by Michel and Kaufmann (1973) at constant temperature of 10° C for 12 h. |
Seed moisture content: To determine amount of imbibed water by primed seeds, a portion of seeds from each Petri dish were weighed just after priming treatments and reweighed after dried, using an oven (75° C, for 48 h). Percentage moisture content was expressed on a fresh weight basis.
Mean germination time: Seeds (25 in each) were placed in Petri dishes between layers of Whatman No. 2 moistened with 10 mL of -10 bar PEG 6000 and distilled water as control. Fungal diseases were controlled by adding 0.15% (a.i.) Captan to the solutions. Two germinators (11 and 18°C) that were regulated at 16 h photoperiod and 80% relative humidity were used. Two millimeters growth of coleoptiles was the criterion for germination. Following formula estimated mean germination time:
![]()
where, Ni is the number of newly germinated seeds at time Ti (Alvarado et al., 1987).
Speed of emergence, seedling vigor index and seedling dry weight. These traits were studied under two temperatures (11 and 18° C) using two separate growth chambers. Considering number of priming treatments (3), water regimes (2) and replications (3), 36 boxes (50 χ30 χ6 cm) containing field soil equally allocated between two growth chambers (18 for each) and again in each growth chamber boxes were divided to two sets of 9 for water stress and non stress treatments. All of the eleven wheat cultivars were sown in each box with 3 cm row space and 2 cm depth. Day and night period were kept 11 and 13 h with 27 and 22° C, respectively. Relative humidity was maintained at 70%. The number of emerged seeds was recorded daily according to the seedling evaluation in the Handbook of AOSA (1990). Water stress started 15 DAS and continued till 25 DAS.
Speed of emergence was calculated using the following formula (Dodlani et al., 1992):
Seedling vigor index was calculated after recording the number of the seedling that had emerged by 5, 7, 9 and 11 DAS and using the following equation (Copeland, 1976): Seedling vigor index = (number of seedling emerged/number of days of first count) + (number of seedling emerged/number of days of second count) + … + (number of seedling emerged/number of days of last count).
Seedling dry weight (mg seedling‾1) of each cultivar was determined by drying normal seedlings obtained at 25 DAS in an oven at 75°C for 48 h.
Statistical analysis: All percentage data were subjected to arcsine transformation before analysis of variance was conducted by the PROC GLM procedure of SAS (1999). Treatment means were considered significantly different using LSD values at a 0.05 probability level.
RESULTS AND DISCUSSION
Amount of imbibed water by hydroprimed and PEG treated seeds across genotypes are demonstrated in Fig. 1. On average, primed seeds with PEG and distilled water imbibed 26.7 and 33.3% water based on fresh weight, respectively. Thus PEG solution significantly reduced seed water imbibition as compared with distilled water which is in confirmation with Kayaa et al. (2006) who found that adverse effect of PEG on seed germination were due to its osmotic effect. This was expected since the lower water potential of the PEG solution should lead to a lower rate of water uptake by seeds. As shown in Fig. 1, seeds of four drought resistant cultivars i.e., Sardari, Agosta-Sefid, Azar 2 and Sabalan imbibed more water than drought susceptible ones in PEG solution indicating the ability of resistant cultivars to absorb more water than susceptible ones in low osmotic potential. Thus, it could be concluded that one possible mechanism of drought resistance is the higher ability of water absorption by germinating seed, especially in unfavorable conditions.
The results of priming treatments and temperature on seedling emergence are shown in Fig. 2. As can be seen, emergence of primed seeds at 11° C with distilled water commenced and maximized a day earlier than non primed seeds which is in line with Dell-Aquila and Tritto (1990) who had reported that primed seeds emerged 12 h earlier than non primed seeds. This may be due to increase in activity of enzymes such as α-amylase, protease and lipase which have great role in break down of macromolecules for growth and development of embryo that ultimately resulted in early and higher seedling emergence. However, although some priming methods enhance germination and emergence in the laboratory and greenhouse, they may have limited practical values for promoting seedling emergence under field condition (Giri and Schillinger, 2003). At both temperatures, hydroprimed and non primed seeds showed the same pattern of seedling emergence with both higher final numbers of emerged seedlings as compared with osmopriming treatment.
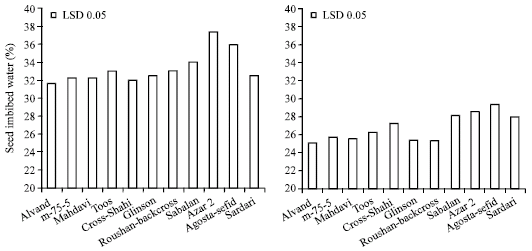 | |
| Fig. 1: | Amount of imbibed water by hydroprimed (left) and PEG treated seeds (right) across genotypes |
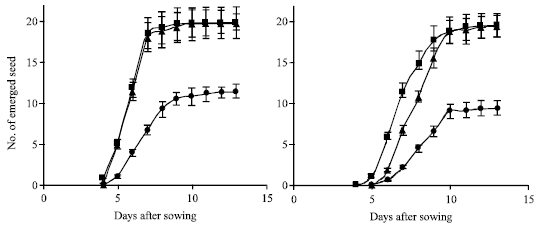 | |
| Fig. 2: | Seedling emergence pattern at 11° C (right) and 18° C (left) across cultivars ( ■) hydroprimed;( ●), primed with PEG and ( ▲) non primed seeds |
Speed of emergence and vigor index of non primed, hydroprimed and PEG treated seeds at 11 and 18° C are shown in Table 1. At 11° C, hydropriming markedly increased speed of emergence and vigor index as compared with non primed seeds, while it had less positive effect on these traits at 18° C. The results of this study imply that hydropriming is more efficient for seedling emergence and establishment at lower temperature which is in confirmation with Akalehiywot and Bewley (1977) who have reported that priming is more suitable for cold area. At both temperatures, seed priming with PEG decreased speed of emergence and vigor index as compared with non primed and hydroprimed seeds. It has been reported by other investigators that osmopriming is less effective method than hydropriming for improving seed germination (Caseiro et al., 2004) and MGT under both salt and drought stress conditions (Kayaa et al., 2006). Although, Bodsworth and Bewley (1981) remarked that -10 bar PEG is a suitable treatment for wheat seed priming, considering the results of present study, seed priming with -10 bar PEG 6000 at 10° C for 12 h is not proposed as a proper treatment for wheat seed enhancement. Superiority of hydropriming on germination may be because, hydroprimed seeds compared to PEG treated seeds were allowed to imbibe more water (Fig. 1). Further, the use of this chemical has some disadvantages such as the reduction of oxygen concentration in the solution due to the viscous nature of PEG which in turn may has negative effects on both protein synthesis and degradation and hamper respiration processes during seed germination.
Regardless of priming treatments and temperatures, drought resistant cultivars i.e., Sardari, Agosta-Sefid, Azar 2 and Sabalan had less speed of emergence and vigor index and more MGT as compared with drought susceptible ones (Table 1). Therefore, drought resistance of wheat cultivars may have no relation with MGT, speed of emergence and seedling vigor index characteristics. It has been reported by other investigators that seed germination indices can not discriminate drought resistant cultivars (Boulsmana and Schapaugh, 1984) and there is no correlation between germination index and crop yield (Hampson and Simpson, 1990). Hence, MGT, speed of emergence and vigor index may be futile for screening of drought resistant genotypes in wheat breeding programs and even cultivar selection based on these indexes may lead to early deletion of drought resistant lines.
Seedling dry weight of hydroprimed and non primed cultivars on 25 DAS at 11 and 18° C under water stress and non stress conditions are shown in Table 2. Seed priming with PEG severely diminished seedling dry weight. Consequently, this treatment was omitted during seedling dry weight evaluation. In general, water stress treatment decreased seedling dry weight across genotypes. Sadeghian and Yavari (2004) reported that seedling growth severely diminished with drought stress and genetic differences were found in sugar beet. At 11° C, hydropriming treatment increased seedling dry weight of 15.3% under water stress condition across genotypes. Thus, increase in seedling dry weight may be one of reasons responsible for positive effects of hydropriming on seed performance under adverse environmental conditions. Nevertheless, reverse results were obtained at 18° C indicating more efficiency of hydropriming in the lower temperature as mentioned before. Regardless of priming treatments and water regimes, seedlings dry weight of drought resistant cultivars were higher than those of susceptible ones at 11° C. Sardari and Azar 2 cultivars had the highest amount of hydroprimed seedling dry weight at 11° C (Table 2). Spra et al. (1991) reported that the higher seedling dry weight, the higher drought resistant capability. Therefore, it is proposed that measurement of seedling dry weight is a proper approach for early screening of wheat drought resistant genotypes which is in conformity with the finding of Boulsmana and Schapaugh (1984) in soybean.
| Table 1: | Mean Germination Time (MGT), speed of emergence and vigor index of non primed, hydroprimed and PEG treated seeds at 11 and 18° C |
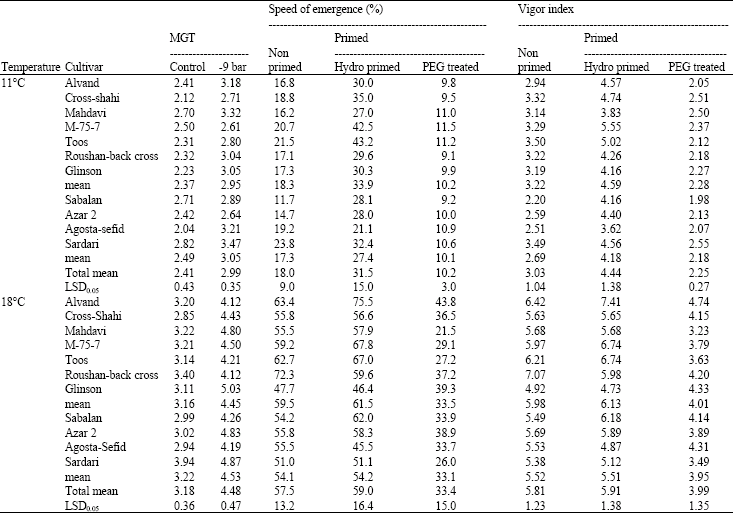 | |
| Table 2: | Seedling dry weight of wheat cultivars (mg seedling‾ 1) on 25 DAS at 11 and 18° C under Water Stress (WS) and non Stress (NS) conditions |
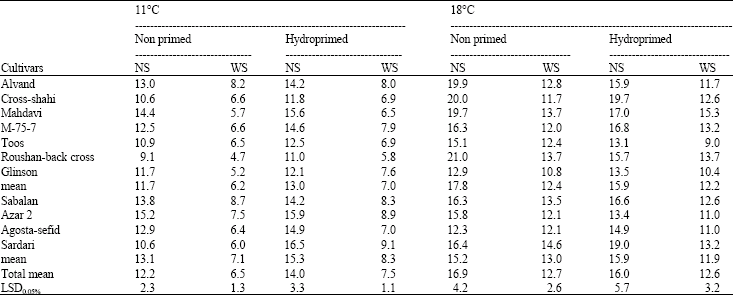 | |
CONCLUSIONS
The data obtained here revealed that hydropriming with distilled water is more efficient than osmopriming with PEG solution for improving MGT, speed of emergence, vigor index and seedling dry weight especially in lower temperature. Lower MGT and enhanced germination index in treated seeds appear to be linked to the efficient production and utilization of germination metabolites and better genetic repair (i.e., earlier and faster synthesis of DNA, RNA and proteins). In the present study, improved seedling dry weight due to hydropriming may be explained by an increased seed water uptake and rate of cell division and earlier start of seedling emergence.
Hydropriming practically ensure rapid and uniform germination accompanied with low abnormal seedling percentage and it has high potential in improving field emergence and ensure early flowering and harvest under stress conditions especially in dry areas. Since its protocol is simple, cheap and dose not requires expansive chemicals and sophisticated equipment, hydropriming can be recommended to farmers to achieve higher germination and uniform emergence, especially in Mediterranean climate in which temperature is low during sowing time.
ACKNOWLEDGMENTS
Authors sincerely appreciate Mehdi Joudi, Vahid Samavati, Rohollah Taghizadah and Kava Lotfi for their valuable help in conducting present research.
REFERENCES
- Alvarado, A.D., K.J. Bradford and J.D. Hewitt, 1987. Osmotic priming of tomato seeds: Effects on germination, field emergence, seedling growth and fruit yield. J. Am. Soc. Hortic. Sci., 112: 427-432.
Direct Link - Bittencourt, M.L.C., D.C.F.S. Dias, L.A.S. Dias and E.F. Araujo, 2005. Germination and vigour of primed asparagus seeds. Sci. Agric., 62: 319-324.
CrossRef - Bodsworth, S. and J.D. Bewley, 1981. Osmotic priming of seeds of crop species with polyethylene glycol as a means of enhancing early and synchronous germination at cool temperatures. Can. J. Bot., 59: 672-676.
Direct Link - Bouslama, M. and W.T. Schapaugh, 1984. Stress tolerance in soybeans. I. Evaluation of three screening techniques for heat and drought tolerance. Crop Sci., 24: 933-937.
CrossRefDirect Link - Caseiro, R., M.A. Bennett and J. Marcos-Filho, 2004. Comparison of three priming techniques for onion seed lots differing in initial seed quality. Seed Sci. Technol., 32: 365-375.
Direct Link - Dell'Aquila, A. and V. Tritto, 1990. Ageing and osmotic priming in wheat seeds: Effects upon certain components of seed quality. Ann. Bot., 65: 21-26.
Direct Link - Dadlani, M., V.V. Shenoy and D.V. Seshu, 1992. Seed coating to improve stand establishment in rice. Seed Sci. Technol., 20: 307-313.
Direct Link - Farooq, M., S.M.A. Basra, M. Khalid, R. Tabassum and T. Mahmood, 2006. Nutrient homeostasis, metabolism of reserves and seedling vigor as affected by seed priming in coarse rice. Can. J. Bot., 84: 1196-1202.
Direct Link - Giri, G.S. and W.F. Schillinger, 2003. Seed priming winter wheat for germination, emergence and yield. Crop Sci., 43: 2135-2141.
CrossRefDirect Link - Hampson, C.R. and G.M. Simpson, 1990. Effects of temperature, salt and osmotic potential on early growth of wheat (Triticum aestivum). I. Germination. Can. J. Bot., 68: 524-528.
CrossRefDirect Link - Harris, D., A.K. Joshi, P. Gothkar and P.S. Sodhi, 1999. On-farm seed priming in semiarid agriculture development and evaluation in maize, rice and chickpea in India using participatory methods. Exp. Agric., 35: 15-29.
Direct Link - Harris, D., 2006. Development and testing of on farm seed priming. Adv. Agron., 90: 129-178.
Direct Link - Hussain, M., M. Farooq, S.M.A. Basra and N. Ahmad, 2006. Influence of seed priming techniques on the seedling establishment, yield and quality of hybrid sunflower. Int. J. Agric. Biol., 8: 14-18.
Direct Link - Kaya, M.D., G. Okcu, M. Atak, Y. Cikili and O. Kolsarici, 2006. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron., 24: 291-295.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Pimentel, D. and M. Pimentel, 2006. Global environmental resources versus world population growth. Ecol. Econ., 59: 195-198.
CrossRefDirect Link - Qin, H. and G.H. Zheng, 1994. Improvement in vigor of hybrid rice seeds and its resistance to imbibition chilling injury. Plant Physiol. Commun., 30: 24-26.
Direct Link - Ruan, S., Q. Xue and K. Tylkowska, 2002. The influence of priming on germination of rice (Oryza sativa L.) seeds and seedling emergence and performance in flooded soil. Seed Sci. Technol., 30: 61-67.
Direct Link - Sadeghian, S.Y. and N. Yavari, 2004. Effect of water-deficit stress on germination and early seedling growth in sugar beet. J. Agron. Crop Sci., 190: 138-144.
CrossRefDirect Link - Mardeh, A.S.S., A. Ahmadi, K. Poustini and V. Mohammadi, 2006. Evaluation of drought resistance indices under various environmental conditions. Field Crop Res., 98: 222-229.
CrossRefDirect Link - Sapra, V.T., E. Savage, A.O. Anaele and C.A. Beyl, 1991. Varietal differences of wheat and triticale to water stress. J. Agron. Crop Sci., 167: 23-28.
CrossRefDirect Link - Basra, S.M.A., M. Farooq, R. Tabassam and N. Ahmad, 2005. Physiological and biochemical aspects of pre-sowing seed treatments in fine rice (Oryza sativa L.). Seed Sci. Technol., 33: 623-628.
CrossRefDirect Link