
Nayer Mohammadkhani
Department of Biology, Faculty of Science, Urmia University, Iran
Reza Heidari
Department of Biology, Faculty of Science, Urmia University, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 4022-4028







ABSTRACT
Water stress is one of the most important environmental factors that reduce growth, development and production of plants. Stress was applied with polyethyleneglycol (PEG) 6000 and water potentials were: zero (control), -0.15 (PEG 10%), -0.49 (PEG 20%), -1.03 (PEG 30%) and -1.76 (PEG 40%) MPa. The roots and leaves respiration of two maize (Zea mays L.) cultivars -704 and 301- were determined in various concentrations of PEG 6000. Oxygen uptake declined in leaves and roots with increasing PEG concentrations. Decrease of oxygen uptake in roots and leaves of 704 variety were higher than 301 variety. Chlorophyll a, b and total chlorophyll content were significantly decreased (p<0.05), but carotenoids content increased (p<0.05) under water stress. Decrease of chlorophyll content in 704 var. was higher than 301 var., but carotenoids content in 301 var. was higher than 704 var. Relative Water Content (RWC) was used to indicate the degree of stress. RWC decreased with increasing PEG concentrations. Lowering of RWC reduced growth and increased shoot/root ratio. Decrease of water content in 704 plants was higher than 301 plants. Shoot/root ratio in 704 var. was higher than 301 var.
PDF Abstract XML References Citation
How to cite this article
Nayer Mohammadkhani and Reza Heidari, 2007. Effects of Water Stress on Respiration, Photosynthetic Pigments and Water Content in Two Maize Cultivars. Pakistan Journal of Biological Sciences, 10: 4022-4028.
DOI: 10.3923/pjbs.2007.4022.4028
URL: https://scialert.net/abstract/?doi=pjbs.2007.4022.4028
DOI: 10.3923/pjbs.2007.4022.4028
URL: https://scialert.net/abstract/?doi=pjbs.2007.4022.4028
INTRODUCTION
Water stress is considered as one of the most important factors limiting plant performance and yield worldwide (Boyer, 1982).
Effects of water stress on plant’s physiology, including growth (MC Donald and Davies, 1996), signaling pathways (Chaves et al., 2003), gene expression (Bray, 2002) and leaf photosynthesis (Flexas et al., 2004a; Lawlor and Cornic, 2002) have been studied extensively.
Surprisingly, compared with other physiological processes, studies examining the effects of water stress on respiration are few (Hsiao, 1973), despite the importance of respiration in ecosystem annual net productivity (Valentini et al., 1999) and the fact that ecosystem respiration is strongly affected by water availability (Bowling et al., 2002).
Respiration rates are often an order of magnitude lower than photosynthesis rates. However, since photosynthesis is limited temporally (i.e., daytime hours) and spatially (i.e., to green biomass), while respiration occurs continuously in every plant organ, the latter may be an equally important factor controlling productivity, particularly when photosynthesis is largely depressed, such as under drought conditions (Flexas et al., 2005; Lawlor and Cronic, 2002).
Under water stress, dehydration of plant tissue can result in an increase in oxidative stress, which causes deterioration in chloroplast structure and an associated loss in chlorophyll. This leads to a decrease in the photosynthetic activity. Chlorophyll content decreased, but carotenoid content increased with increase of PEG concentrations.
The relative water content in different maize cultivars decreased significantly with drought stress (Chen and Dai, 1994; Lu and He, 1995). Furthermore water stress decreased the relative water content in seedlings of a drought-sensitive cultivars (Li and van staden, 1998b). Song et al. (1995) also reported that maize leaves with drought-tolerant cultivars had relatively high water content.
Polyethylene glycol (PEG) 6000 is described as a non-ionic water-soluble polymer, which is not expected to penetrate plant tissues rapidly and is widely used to induce water stress in higher plants (Nepomuceno et al., 1998). There is no evidence that PEG is harmful to maize seedling roots (Verslues et al., 1998).
The aim of the present study was to undertake a comparative analysis of the effects of water stress on respiration rates in roots and shoots in two maize cultivars and their relation to photosynthetic pigments variations. Some specific questions were addressed:
| • | Which are the changes in respiration rates in roots and leaves in two cultivars? |
| • | Which are the changes in photosynthetic pigments content in leaves of two cultivars? |
| • | Is respiration rate affected by a decrease in water availability? |
MATERIALS AND METHODS
Plant materials and growth conditions: This study was conducted at biochemistry Laboratory, Department of Biology, Urmia University, Iran, during the winter of 2007.
Two genotypes of maize (Zea mays L.) -var. 704 and var. 301- were used. The seeds of both cultivars were germinated in Petri dishes on two layers of filter paper at 25°C in an incubator. After three days, the seedlings transferred to plastic pots (15 cm diameter, 20 cm depth) filled with sand and irrigated with half strength of Hoagland nutrient solution.
Six-days seedlings were removed from the sand, washed with tap water, dried and transferred to hydroponics culture of aerated test tubes containing polyethylene glycol (PEG) 6000 solutions of 10, 20, 30 and 40% strengths to achieve water deficit levels of -0.15, -0.4.9, -1.03 and -1.76 MPa, respectively (Burlyn et al., 1973; Nicholas, 1989; Steuter et al., 1981) as treatments and aerated test tubes containing half strength Hoagland nutrient solution which served as control.
Stress was applied for 24 h and then respiration rate, photosynthetic pigments content and relative water content were determined.
Respiratory measurements: Oxygen uptake of roots and leaves were measured at 25°C using an oxygen meter (WTW model oxi 730). Roots and leaves segments (approximately 0.5 g fresh weight) were placed in 4 mL reaction medium [0.25 M sucrose, 0.01 M Tris, 0.01 M K2HPO4, 0.005 M MgCl2, 0.005 M EDTA, 0.5 mg mL‾1 BSA] adjusted to pH = 7.2 with HCl and O2 measured in period of 2 min (John and James, 1970).
Measurement of photosynthetic pigments content: The chlorophylls and carotenoids (carotene and xanthophylls) content of leaves measured with Lichtenthaler and Wellbum (1983) method. The pigments of 0.1 g of leaf fresh weight extracted by acetone 80%. Extracts filtered by filter paper absorbance of samples was measured at 663.2, 646.8 and 470 nm by Uv-visible spectrophotometer (WPA model S2100).
Chlorophyll a, b, total chlorophyll and carotenoids content were measured with following equations:
| Chlorophyll a | = | 12.25 A 663.2-2.798 A 646.8 |
| Chlorophyll b | = | 21.5 A 646.8-5.1 A 663.2 |
| Total chlorophyll | = | chlorophyll a + chlorophyll b |
| Carotenoids content | = | (1000 A470-1.82 chlorophyll a-85.02 chlorophyll b)/198 |
Plants water status: Relative water content was determined with following equation:
![]()
Fresh weight of the plants was measured and after that plants were dried at 105°C until reached constant weight for the determination of dry weight. To determine the turgid weight, samples were soaked in distilled water for 4 hours at room temperature (approximately 20°C) and then turgid weight was measured (Fletcher et al., 1988).
Statistical analysis: Mean values were taken from measurements of four replicates and Standard Error of the means was calculated.
Differences between means were determined by One-way ANOVA and Turkey’s multiple range tests (p<0.05). Analyses were done using the Statistical Package for Social Sciences (SPSS) for windows (version 13.0).
RESULTS
Effects of water stress on respiration rate: Lowering their water potential somatically retarded the uptake of O2 by root and leaf segments. The changes in leaf and root respiratory rate could result from damage to the mitochondria themselves or in leaf which altered substrate availability due to inhibition of photosynthesis.
Leaf respiration averaged 17.22 μmol O2 g‾1 Fw min‾1 in control plants, 704 var. and 14.46 μmol O2 g‾1 Fw min‾1 in 301 var. Under severe water stress (PEG 40%) respiration was lower 704 var. than 301 var., although not significantly different (6.22 μmol O2 g‾1 Fw min‾1 in 704 var. and 7.06 μmol O2 g‾1 Fw min‾1 in 301var.).
In roots of control plants, respiration averaged 10.98 μmol O2 g‾1 Fw min‾1 in 704 var. and 9.06 μmol O2 g‾1 Fw min‾1 in 301var. Under severe stress (PEG40%) respiration was lower than control (4.17 μmol O2 g‾1 Fw min‾1 in 704 var. and 5.57 μmol O2 g‾1 Fw min‾1 in 301var).
Therefore, respiration decreased with increasing PEG treatments (Fig. 1). In PEG 10%, leaf respiration was 0.76 fold in 704 var. and 0.77 fold in 301 var. as compared to control plants and in PEG 40%, leaf respiration was 0.36 fold in 704 plants and 0.49 fold in 301 plants as compared to control plants.
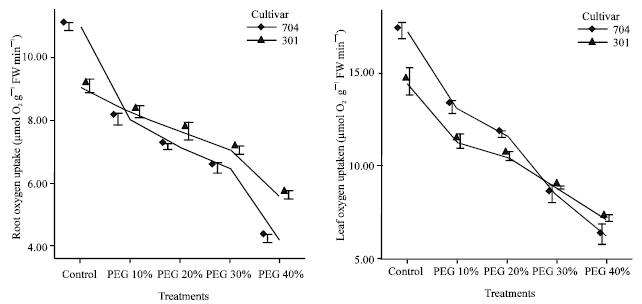 | |
| Fig. 1: | Effects of different PEG 6000 concentrations on oxygen uptake in roots and leaves of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
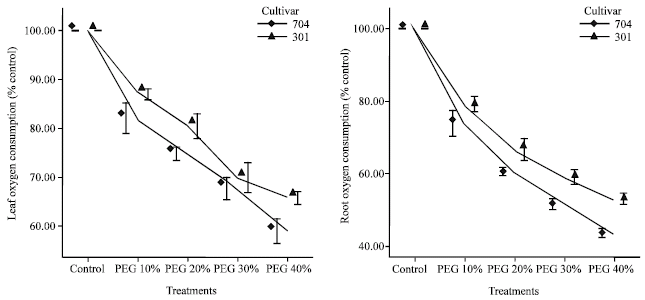 | |
| Fig. 2: | Effects of different PEG 6000 concentrations on oxygen consumption percent in roots and leaves of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
About root respiration, in PEG 10%, this factor was 0.73 fold in 704 var. and 0.91 fold in 301 var. and in PEG 40%, root respiration was 0.38 fold in 704 var. and 0.62 fold in 301 var. as compared to control plants. The decrease of respiration rates in roots and leaves in 704 var. were higher than 301 var.
The percentage of oxygen consumption decreased gradually with increasing PEG concentrations (Fig. 2). The percentage of oxygen consumption in PEG 40% (water potential -1.76 Mpa), in roots decreased to 43.63% in 704 var. and 52.98% in 301 var. and in leaves decreased to 58.8% in 704 var. and 65.85% in 301 var.
The decrease of oxygen consumption in 704 var. was higher than 301 var. and in roots was higher than leaves.
Effects of water stress on photosynthetic pigments: Drought stress caused a reduction in chlorophyll a, b and total chlorophyll content in both varieties, but the decrease in 704 var. was higher than 301 var.
In PEG 40%, chlorophyll a content was 0.41 fold in 704 var. and 0.40 fold in 301 var. as compared to control plants. In highest water stress, chlorophyll b content was 0.20 fold in 704 var. and 0.47 fold in 301 var. as compared to control plants and the decrease in 301 var. was gradually and in 704 var. in PEG 40% was enormous.
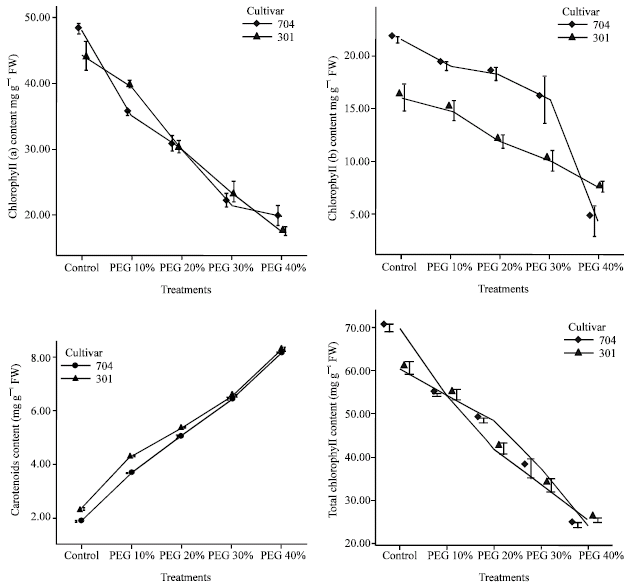 | |
| Fig. 3: | Effects of different PEG 6000 concentrations on photosynthetic pigments in leaves of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
Total chlorophyll content decreased in both varieties and in PEG 40%, this factor was 0.35 fold in 704 var. and 0.42 fold in 301 var. as compared to control (Fig. 3).
Carotenoids content in both varieties increased, but the increase in 301 var. was higher than 704 var. In PEG 40%, carotenoids content was 4.39 fold in 704 var. and 3.97 fold in 301 var. as compared to control plants. It means that 301 plants have higher carotenoids content and lower chlorophyll content than 704 plants when water stressed.
Effects of water stress on relative water content: The RWC of leaves and roots indicated the extent of dehydration. In roots and shoots in control plants, 704 var. have higher water content than 301 var., but in severe water stress (water potential -1.76 MPa), 301 plants roots and shoots have higher water content than 704 plants (Fig. 4).
Relative water content in roots and shoots decreased with increasing PEG 6000 concentrations. IN PEG 40%, this decrease was enormous in roots and shoots in both varieties. In PEG 40%, RWC was 0.41 fold in 704 var. and 0.62 fold in 301 var. in roots and 0.53 fold in 704 var. and 0.61 fold in 301 var. in shoots as compared to control plants.
With increase of PEG concentrations, the shoot/root ratio was increased and shoot/root ratio in 704 plants was higher than 301 plants (Fig. 5). In PEG 40%, shoot/root ratio was 1.29 fold in 704 plants and 1.12 fold in 301 plants as compared to control.
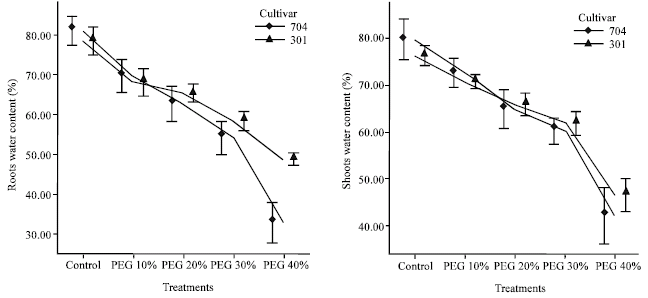 | |
| Fig. 4: | Effects of different PEG 6000 concentrations on water content in roots and shoots of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
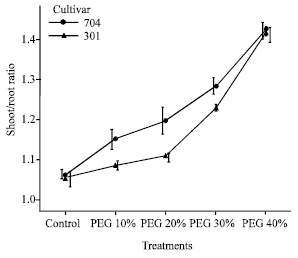 | |
| Fig. 5: | Effects of different PEG 6000 concentrations on shoot/root ratio of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
DISCUSSION
Increasing PEG concentration clearly depressed both roots and shoots respiration. The addition of PEG is responsible for declining water potential and osmotic withdrawal or water.
While several studies described water stress induced decreased respiration rate (Brix, 1962; Brown and Thomas, 1980; Palta and Nobel, 1989; Gonzalez-Meler et al., 1997; Ghashghaire et al., 2002; Haupt-Herting et al., 2001), others have shown unaffected rates (Lawlor, 1976; Loboda, 1993).
Leaf respiration rates decreased under drought in most species, but the decline was always smaller than that of photosynthesis, therefore resulting in decreased photosynthesis-to respiration ratio (indicative of leaf carbon balance).
The decline in respiration in response to drought seems to be part of a systemic metabolic response, which occurs under conditions where drought severely restricts CO2 availability inside leaf cells, therefore, creating the risk of a secondary oxidative stress (Flexas et al., 2004a,b, 2006).
Our original objective from this study was finding the changes in respiration rates in leaves and roots with increasing PEG treatments. We found that with increase of drought stress, respiration rates decreased in both roots and leaves. These results supported previous findings (Brix, 1962; Brown and Thomas, 1980; Palta and Nobel, 1989; Gonzalez-Meler et al., 1997; Ghashghaire et al., 2002; Haupt-Herting et al., 2001). In PEG 40%, respiration rate in 704 plants in roots and leaves were not only lower than the control, but were also lower than in 301 var. It means that in leaves and roots the decrease of oxygen uptake in 704 var. was higher than 301 var.
With increase of PEG concentrations, oxygen consumption percentage decreased in both roots and leaves. The decrease of oxygen consumption percentage in 704 plants was higher than 301 plants. Therefore, water stress has a higher effect in 704 plant’s respiration than 301 plants. Oxygen consumption percentage in high water stress in roots was lower than leaves and roots were more sensitive than leaves. Drought probably acted directly on roots, because the roots were immerged in PEG solutions and water stress in roots was higher than shoots, whereas leaves could reduce oxygen consumption by stoma regulations.
There were a positive and strong correlation between respiration rates and water content. The decreased respiration rate showed positively correlated to decrease of relative water content. Respiration rates affected by a decrease in water content. It means that a decrease in water content caused a decrease in oxygen consumption and respiration rates (Brix, 1962; Crafts, 1968). The decrease of water content in 704 plants was higher than 301 plants and in roots was higher than shoots. Three hundred and one plants have higher water content than 704 plants in roots and shoots in both varieties.
The shoot/root ratio was increased in water stress and this factor in 704 var. was higher than 301 var.
In water stress, with increasing PEG treatments, chlorophyll a, b and total chlorophyll content decreased. In 704 plants chlorophyll content in leaves decreased higher than 301 plants, but in severe water stress, 704 plants have higher total chlorophyll content than 301 plants. Increase of carotenoids content in 301 var. was higher than 704 plants. Therefore 301 plants have a better protection than 704 plants, because carotenoids have a protective role and protect chlorophyll from photo oxidation.
The above results suggest that plants of the 301 variety have a better tolerance to water stress as compared to 704 variety.
It suggest that effect of water stress on respiration rates and its relations to relative water content and photosynthesis rates study in other varieties of maize and in other plants, especially correlation between respiration rates and photosynthesis rates in severe water stress.
ACKNOWLEDGMENTS
This study supported by Department of Biology of Urmia University, Iran. We greatly acknowledge Dr. M. Ilkhanipour for her support in providing facilities. We greatly acknowledge Prof. Khalafi for modifying this manuscript.
REFERENCES
- Boyer, J.S., 1982. Plant productivity and environment. Science, 218: 443-448.
CrossRefPubMedDirect Link - Bray, E.A., 2002. Abscisic acid regulation of gene expression during water-deficit stress in the era of the Arabidopsis genome. Plant Cell Environ., 25: 153-161.
CrossRefPubMedDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Chaves, M.M., J.P. Maroco and J.S. Pereira, 2003. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol., 30: 239-264.
CrossRefDirect Link - Chen, J. and J.Y. Dai, 1994. Correlation among photosynthesis, lipid per oxidation ultra structural changes of mesophyll cells in corn leaves under water stress. Maize Sci., 2: 36-40.
Direct Link - Fletcher, R.A., M. Santakumari and D.P. Murr, 1988. Imposition of water stress in wheat seedlings improves the efficiency of uniconazole induced thermal resistance. Physiol. Plant., 47: 360-364.
CrossRefDirect Link - Flexas, J., J. Bota, F. Loreto, G. Cornic and T.D. Sharkey, 2004. Diffusive and metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant Biol., 6: 269-279.
CrossRefDirect Link - Flexas, J., J. Bota, J. Cifre, J.M. Escalona and J. Galmes et al., 2004. Understanding down-regulation of photosynthesis under water stress: Future prospects and searching for physiological tools for irrigation management. Ann. Applied Biol., 144: 273-283.
Direct Link - Flexas, J., J. Bota, J. Galmes, H. Medrano and M. Ribas-Carbo, 2006. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant., 127: 343-352.
CrossRefDirect Link - Gonzalez-Meler, M.A., 1997. Effects of prolonged drought stress and nitrogen deficiency on respiratory O2 uptake of bean and pepper leaves. Photosynthetica, 34: 505-512.
Direct Link - Haupt-Herting, S., 2001. A new approach to measure gross CO2 fluxes in leaves. Gross CO2 assimilation, photo respiration and mitochondrial respiration in the light in tomato under drought stress. Plant Physiol., 126: 388-396.
Direct Link - Hsiao, T.C., 1973. Plant responses to water stress. Annu. Rev. Plant Physiol., 24: 519-570.
CrossRefDirect Link - Lawlor, D.W. and G. Cornic, 2002. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ., 25: 275-294.
CrossRefDirect Link - Li, L. and J. van Staden, 1998. Effects of plant growth regulators on drought resistance of two maize cultivars. S. Afr. J. Bot., 64: 116-120.
CrossRefDirect Link - Lichtenthaler, H.K. and A.R. Wellburn, 1983. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Loboda, T., 1993. Gas exchange of different spring cereal genotypes under normal and drought conditions. Photosynthetica, 29: 567-572.
Direct Link - Nepomuceno, A.L., D.M. Oostrerhuis and J.M. Stewart, 1998. Physiological responses of cotton leaves and roots to water deficit induced by Polythylene. Glycol. Environ. Exp. Bot., 40: 29-41.
Direct Link - Money, N.P., 1989. Osmotic pressure of aqueous polyethylene glycols: Relationship between molecular weight and vapor pressure deficit. Plant Physiol., 91: 766-769.
CrossRefDirect Link - Steuter, A.A., A. Mozafar and J. Goodin, 1981. Water potential of aqueous polyethylene glycol. Plant Physiol., 67: 64-67.
Direct Link - Valentini, R., G. Matteucci, A.J. Dolman, E.D. Schulze and C. Rebmann et al., 2000. Respiration at the main determinant of carbon balance in European forests. Nature, 404: 861-865.
CrossRef - Verslues, P.E., E.S. Ober and R.E. Sharp, 1998. Root growth and oxygen relations at low water potentials. Impact of oxygen availability in polyethylene glycol solutions. Plant Physiol., 116: 1403-1412.
PubMed