
A. Mehrvar
Department of Plant Protection, Maragheh Higher Education Complex, University of Tabriz, Tabriz-5166616471, Iran
R. J. Rabindra
Project Directorate of Biological Control (ICAR), P.B. No. 2491, Bellary Road, Hebbal, Bangalore-560024, India
K. Veenakumari
Project Directorate of Biological Control (ICAR), P.B. No. 2491, Bellary Road, Hebbal, Bangalore-560024, India
G. B. Narabenchi
Pest Control (India) (PCI), Bio-Control Research Laboratories (BCRL),
Bangalore-560064, India
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 22 | Page No.: 3992-3999







ABSTRACT
The effect of four major parameters, viz., larval age and weight, inoculation dose, incubation temperature and time of harvesting the larvae, on the production of three isolates of nucleopolyhedrovirus of Helicoverpa armigera (HaNPV), viz., Ooty (OTY), Coimbatore (CMB) and Negamum (NGM) were evaluated. Early 5th instar larvae recorded the maximum yield of virus per larva when inoculated with a dose of 5χ105 POB larva‾1 and incubated at a temperature of 25°C. Also, highest POB yield was recorded when virosed larvae were harvested as cadavers. However, among the isolates tested in this study, CMB isolate collected from Tamil Nadu, India, showed the highest yield per larva in all of the conditions.
PDF Abstract XML References Citation
How to cite this article
A. Mehrvar, R. J. Rabindra, K. Veenakumari and G. B. Narabenchi, 2007. Standardization of Mass Production in Three Isolates of Nucleopolyhedrovirus of Helicoverpa armigera (Hübner). Pakistan Journal of Biological Sciences, 10: 3992-3999.
DOI: 10.3923/pjbs.2007.3992.3999
URL: https://scialert.net/abstract/?doi=pjbs.2007.3992.3999
DOI: 10.3923/pjbs.2007.3992.3999
URL: https://scialert.net/abstract/?doi=pjbs.2007.3992.3999
INTRODUCTION
Since viral pathogens are obligate in nature, they must be necessarily multiplied on their natural live hosts from which they have been collected (Narayana, 2003). The selection of a virus for pest management depends not only on its bioefficacy but also on the ease of mass production (Sherman, 1985). Nucleopolyhedroviruses find extensive use in assisted control programmes for early management of pests (Carner and Yearian, 1989; Rabindra et al., 2003; Dolinski and Lacey, 2007). While propagation of the virus in cell culture continues to receive increasing attention (Shuler et al., 1995), in vivo production has been found to be the only feasible method for large scale propagation of virus (Hunter et al., 1998). There has been continuous flow of information on the in vivo production of baculoviruses, notably the nuclear polyhedrosis viruses in many insects (Shapiro, 1986; Ebora et al., 1990). However, to increase the yield of three HaNPV isolates collected from India and selection the highly promising viral isolate under in vivo conditions a series of laboratorial studies were conducted. The present investigations were therefore undertaken to evaluate the effect of stage of the host larvae, incubation temperature and dose of virus inoculum as well as the time of harvest on the in vivo yield of three HaNPV isolates.
MATERIALS AND METHODS
The H. armigera culture used in the study was maintained on a semi-synthetic diet based on Shorey and Hale (1965) in Bio-Control Research Laboratories (BCRL), Bangalore, India. Three Indian isolates of HaNPV collected from Coimbatore (CMB), Negamum (NGM) and Ooty (OTY) of Tamil Nadu state, India, were used in this study during the year 2005. These isolates were passaged through early fifth instar larvae of host insect at 25±1°C to get uniformity in their virulence since they had been stored in the refrigerator for prolonged period of time. All the experiments were performed in insect virology laboratory of Project Directorate of Biological Control (PDBC), Bangalore, India.
Effect of larval age and weight on virus yield: Mid 4th, late 4th, early 5th and mid 5th instar larvae of H. armigera were used to study the influence of insect age on virus yield. Semi-synthetic diet without formaldehyde was prepared and filled in 5 mL glass vials up to 1/3rd height of the vial. A quantity of 10 μL of virus suspension (5χ105 POB larva‾1) was applied onto the diet surface using a micropipette providing a dose of 1965.78 POB mm‾2. The suspension was spread uniformly over the diet surface with the polished blunt end of a glass rod (6 mm). Larvae in the four age groups were weighed individually in an electronic balance and transferred to treated diet. The treatments were replicated three times and included a control for each age group and isolate. Each treatment had 40 insects. After inoculation, the larvae were incubated at 25±1°C in an incubator. The mortality in different treatments was recorded at 24 h interval. Upon death, the cadavers were collected, transferred to sterile vials and the number of harvestable cadavers in each treatment was recorded. The samples were frozen immediately.
The cadavers were homogenized individually using a glass pestle and mortar with distilled water. The homogenate was transferred to a measuring cylinder and volume made up to 25 mL with distilled water. Enumeration of polyhedra was performed in this stage and the following parameters were calculated:
![]()
Yield per 100 inoculated larvae (POB) = Yield larva‾1 χ Corrected larval mortality (%)
![]()
Effect of inoculation dose on virus yield: The effect of inoculation dose on the yield of the virus was studied against early 5th instar larvae. The larvae were exposed to different doses of 1965.87, 393.17 and 78.63 POB mm‾2 of the isolates. Aliquot of 10 μL virus suspension was applied on the diet surface as mentioned earlier. Each treatment had 40 larvae in three replications for each isolate. After inoculation the larvae were incubated at 25±1°C in an incubator. Untreated controls for the respective isolates and doses were maintained similarly. The mortality was recorded at 24 h interval and the experiment was terminated on 10th day. The cadavers were collected daily, frozen and processed and yield was assessed as described earlier.
Effect of inoculation temperature on virus yield: Early 5th instar larvae were exposed to 1965.87 POB mm‾2 of diet surface of each viral isolate. Similar procedure was followed as described above except the incubation temperatures were either room temperature (28-31°C), 25±1°C, or 30±1°C.
Effect of period of larval harvest on virus yield: Early 5th instar larvae of H. armigera were allowed to feed on diet surface treated with viral dose of 1965.87 POB mm‾2 and incubated at 25±1°C for each NPV isolate. The infected larvae were harvested five, six, seven and eight days after inoculation. Also, a treatment of harvesting the cadavers was included for comparison. An untreated control was also maintained. Each treatment was replicated three times with 40 larvae per replication.
The larval cadavers were transferred to sterile vials, plugged with cotton and frozen immediately. The cadavers were then processed and POBs were enumerated as described earlier.
Statistical analyses: Analyses of variances (ANOVA) were carried out using SAS software version 6.12 and means were separated by Duncan’s Multiple Range Test (DMRT). All data in percentage were transformed to arcsin √percentage and the data from POB yields were subjected to log transformation and then analyzed. The larval counts were also transformed to √x+0.5 values. The probit analyses in various experiments were carried out in a Statistical Package for Social Sciences (SPSS), version 10.0 for windows.
RESULTS
Virus yield in relation to larval age and weight: A significant influence of larval age and weight on the per cent larval mortality, yield larva‾1, yield per 100 inoculated larvae and productivity ratio of the isolates was observed in the assay (Table 1). Of the different larval ages, early 5th instar recorded significantly the highest yield and productivity with all the three isolates tested. Mid 4th, late 4th and mid 5th instars registered decreased production of POB.
The initial larval weight was critical for obtaining higher POB yield in all the isolates tested in this study. The maximum POB yield was related to the early 5th instar larvae with initial weight range of 65.46-68.13 mg (Fig. 1).
Virus yield in relation to inoculation dose: Optimization studies on inoculation dose revealed that larval mortality increased as the inoculum dose advanced. Highest mortality of 83.77, 82.90 and 82.05% was recorded at inoculum dose of 1965.87 POB mm‾2 of NGM, OTY and CMB isolates, respectively (Table 2). Similarly, the virus yield per larva and for 100 inoculated larvae was significantly higher at this dose of all the three isolates. The productivity ratio of all the isolates decreased progressively as the dose of inoculation advanced.
| Table 1: | Effect of larval stage on the yield of HaNPV isolates at an inoculation dose of 5χ105 POB larva‾1 and an incubation temperature of 25±1°C |
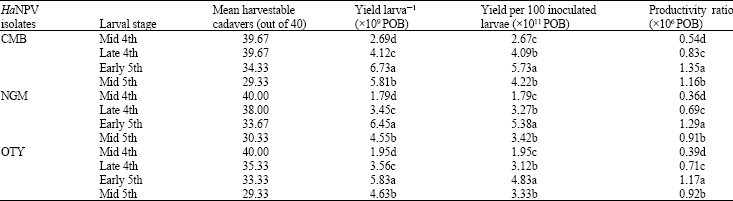 | |
| In a column, for each isolate means followed by the same letter (a-d) are not significantly different (p = 0.05) by Duncan’s Multiple Range Test (DMRT) | |
| Table 2: | Effect of dose of inoculum on the yield of HaNPV isolates achieved from early 5th instar larvae at an incubation temperature of 25±1°C |
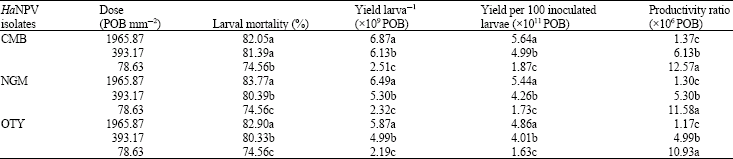 | |
| In a column, for each isolate means followed by the same letter (a-c) are not significantly different (p = 0.05) by DMRT | |
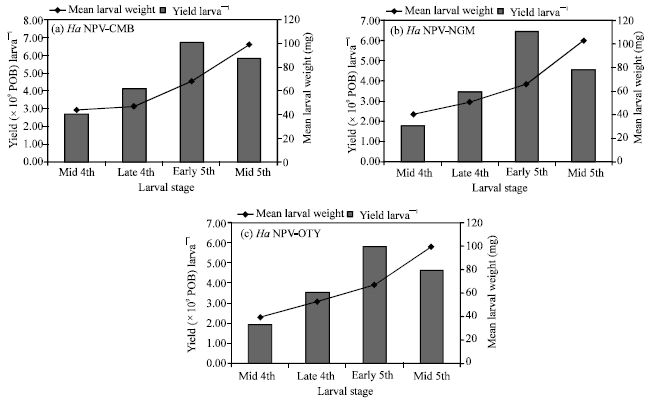 | |
| Fig. 1: | Effect of larval stage and weight on the yield of Ha NPV isolates at an inoculation dose of 5χ105 POB larva‾1 and an incubation temperature of 25±1°C |
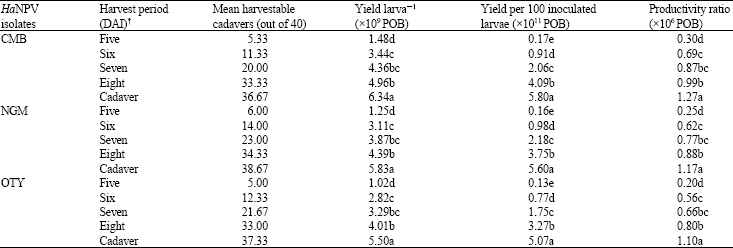
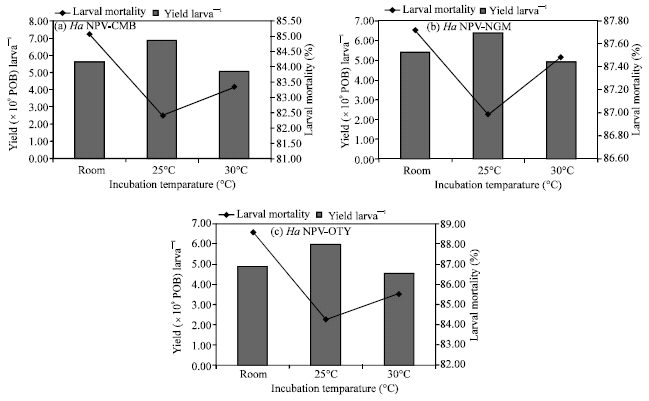
Virus yield in relation to incubation temperature: The maximum yield of virus per larva and yield per 100 inoculated larvae were obtained for all the viral isolates when larvae were incubated at 25°C after virus inoculation as compared with that of room temperature and 30°C (Table 3).
| Table 3: | Effect of incubation temperature on the yield of HaNPV isolates achieved from early 5th instar larvae inoculated with 5χ105 POB larva‾1 |
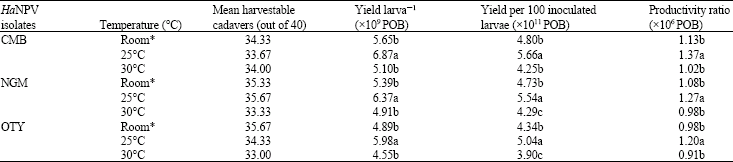 | |
| *: The temperature ranged 28-31°C during the period of study. In a column, for each isolate means followed by the same letter (a-c) are not significantly different (p = 0.05) by DMRT | |
| Table 4: | Effect of harvest period on the yield of HaNPV isolates achieved from early 5th instar larvae inoculated with 5χ105 POB larva‾1 at an incubation temperature of 25±1°C |
 | |
| ♣: Days after inoculation. In a column, for each isolate means followed by the same letter (a-e) are not significantly different (p = 0.05) by DMRT | |
 | |
| Fig. 2: | Effect of incubation temperature on the yield of HaNPV isolates and larval mortality achieved from early 5th instar larvae inoculated with 5χ105 POB larva‾1 |
 | |
| Fig. 3: | Effect of harvest period on the yield of HaNPV isolates and larval mortality achieved from early 5th instar larvae inoculated with 5χ105 POB larva‾1 at 25°C |
The productivity was also the highest at 25°C. The yield larva‾1, yield per 100 inoculated larvae and productivity ratio were minimum at 30°C.
Data on the larval mortality showed a rise in incubation temperature resulted in a significant increase in the per cent kill in all the isolates (Fig. 2).
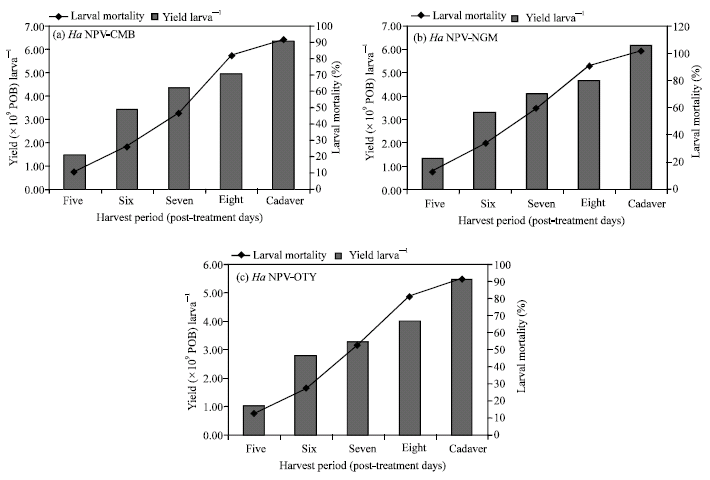
Effect of periods of larval harvest on virus yield: There was a significant increase in larval mortality with increasing time of larval harvest in all the three viral isolates (Table 4). The yield larva‾1, yield per 100 inoculated larvae and productivity ratio were the highest when virosed larvae were harvested as cadavers. As the time of harvest decreased, POB yield larva‾1, yield per 100 inoculated larvae and productivity ratio were reduced (Fig. 3).
DISCUSSION
Virus yield in relation to larval age and weight: The yield and productivity parameters of the viral isolates were determined by larval age and weight at the time of inoculation. Of the different age groups studied, early 5th instar with a weight range of 65.46-68.13 mg recorded significantly the highest yield of 6.73χ109, 6.45χ109 and 5.83χ109 POB larva‾1 in the case of CMB, NGM and OTY isolates, respectively. Mid 4th, late 4th and mid 5th instar larvae registered lower POB production (Table 1). Teakle and Byrne (1989) found an exponential increase in the yield of the virus with age of the larvae of H. armigera. A 100-fold increase in the yield of the virus was noted in larvae of age 6 days compared to one day old larva. Ignoffo (1966) estimated that at least 6χ109 POB were produced per larva in later instars of H. zea. Dhandapani (1990) reported in a study on H. armigera that the yield of POB larva‾1 inoculated at the 5th instar stage was 2.40χ109. Early 5th S. litura larvae was found to be an optimum stage for inoculation of virus to maximize the virus productivity (Tuan et al., 1998; Subramanian et al., 2001). Monobrullah and Nagata (2000) reported that 9-day-old S. litura larvae weighing 125-155 mg treated with 4.8χ106 POB larva‾1 through diet resulted in maximum productivity of the NPV. Shieh (1989) reported that from H. zea larvae with the initial larval weight of 50-120 mg, the highest POB yields could be obtained. At this weight range, the larvae could continue their normal growth as that of healthy insects until a day before death resulting in a higher amount of POB yields. These observations are in agreement with the present findings. When the larvae of H. armigera were allowed to feed on a constant dose, Whitlock (1977) found an inverse relationship between larval age at the time of inoculation and the mortality. In the present studies, mortality was significantly highest in the 4th and mid 4th instar larvae followed by early 5th and mid 5th instars. However, in all the HaNPV isolates tested, yield per 100 inoculated larvae was the highest in the early 5th instar (Table 1).
Virus yield in relation to inoculation dose: A significant influence of inoculation dose on the per cent larval mortality and yield larva‾1 was observed with all the isolates tested. The highest mortality occurred with the highest inoculation dose of 1965.87 POB mm‾2, recording 82.05, 83.77 and 82.90% for the isolates CMB, NGM, OTY, respectively. These results are in agreement with the findings of Narayanan and Jayaraj (2002) who observed a marked difference between doses as well as a significant interaction between dosages and larval instars. A mean number of 2.5χ1010 POB was harvested from late 4th instars when inoculated at low dose of 1.1χ104 POB cup‾1 compared to 1.2χ1010 POB when inoculated at a higher dose of 1.1χ106. A virus concentration of 3χ106 POB mL‾1 by diet incorporation technique (Tuan et al., 1998) and 1χ108 POB mL‾1 by diet surface contamination method (Subramaninan et al., 2001; Kumar and Rabindra, 2003) were found to be optimum for in vivo production of the virus. However, the productivity ratio progressively decreased as the dose of inoculation increased. This is in partial agreement with the findings by Bell (1991) who reported that a lower dose of virus can be used for achieving higher yield.
Virus yield in relation to incubation temperature: Data on evaluation of different incubation temperature on the yield of three HaNPV isolates indicated that, maximum yield larva‾1 was obtained at 25°C followed by room temperature and 30°C (Table 3). Similarly, the yield per 100 inoculated larvae and productivity was also maximum at 25°C. However, the maximum larval mortality occurred at room temperature followed by 30 and 25°C (Fig. 2).
Insect systems function optimally within a limited range of temperature (Chapman, 1998). Synchrony in larval development during virus multiplication is considered important, as the larvae are sensitive to temperature fluctuation. Any variation in optimal temperature will directly affect the larval growth as well as viral multiplication. So, handling this synchronization to get the maximum benefit in terms of yield and productivity of the virus was one of the major objectives in the present studies. Optimization of larval growth during the virus incubation will allow the virus to attain rate of its multiplication. Studies conducted by O’Reilly and Miller (1989) indicated that prolongation in larval growth, even beyond the period of a normal larval stage, would benefit the viral reproduction. Present findings are in agreement with that of previous studies.
Cherry et al. (1997) indicated that the productivity was maximum at an incubation temperature of 25°C for S. litura and S. exempta. Huang (1995) reported that a temperature range of 24-27°C was optimum for 3rd and 4th instar S. litura larvae inoculated with 3.85χ105 and 3.85χ107 POB mL‾1, respectively, to maximize the SlNPV yield. Incubation of early 5th instar S. litura larvae dosed with 3932.4 POB mm‾2 at 25°C enhanced the NPV productivity to 6.623χ1011 POB yield per 100 inoculated larvae, while it was only 1.779χ1011 at 35°C (Subramanian et al., 2006).
Many laboratory studies have demonstrated that nucleopolyhedrovirus is inactivated by exposure to high temperature. McLeod et al. (1977) stated that increase in temperature from 15 to 45°C increased the LD50 values of H. zea NPV (29.8-349.2 POB mm‾2 of diet surface). Stairs (1978) indicated that high temperatures caused direct inactivation of the virus and adversely affected the viral replication. Johnson et al. (1982) demonstrated the inhibition of virus activity against the velvet bean caterpillar, Anticarsia gemmatalis, at the extremes of temperature of 10 and 40°C. Kelly and Entwistle (1988) found an approximate linear relationship between the Mamestra brassicae NPV and the incubation temperature. Histopathological studies by Sathiah (2001) revealed that at 25°C, the growth of fat body in virus-inoculated larvae progressed normally during the early stages of infestation providing adequate substrate for the growth and multiplication of the virus. In H. armigera larvae, the virus multiplied at a slow pace at 25°C allowing the fat bodies to proliferate simultaneously. At higher temperatures the virus multiplied faster destroying the fat body before it could grow to provide greater substrate volume. Therefore, a good mass production facility should possess a temperature-controlled incubation chamber to provide a constant temperature of 25±1°C.
Virus yield in relation to period of harvest: Harvesting of virosed larvae is usually considered as a laborious and time consuming job in all the virus production units. Therefore attempts were made to optimize the period of harvest as it has influence on biological activity of the virus and growth of secondary contaminants like bacteria. The POB yield was higher when virosed larvae were harvested as cadavers than when harvested at different days after inoculation (Table 4). The POB yield per 100 inoculated larvae and productivity ratio were also higher in the case of all isolates when harvested from cadavers. The finding of this study is in contrast with that of Smith and Vlak (1988) who reported that production of S. exigua NPV did not increase after seven days of inoculation. However, Sathiah (2001) and Narabenchi (2004) state that harvesting the virosed larvae as cadavers will enhance the yield production of HaNPV.
Ignoffo and Shapiro (1978) found that the activity of H. zea NPV processed from dead hosts was 7-9 times higher than those from live virosed larvae. Similarly, Shapiro and Bell (1981) evaluated the yield and biological activity of Lymantria dispar NPV at different times after infection. They showed that POB harvested from dead larvae were up to 7 times more active than from living infected larvae. Also, the virus yield increased up to about 2χ109 POB larva‾1 during the first 11 days and then remained constant.
CONCLUSIONS
Virus mass production in vivo in host larvae is the pragmatic method as of today. A variety of parameters have been addressed for increasing productivity of the HaNPV. Present study revealed that factors like larval age and weight, dose of the inoculum, the incubation temperature and the period of harvest could enhance the in vivo yield of the virus. Of the larval age evaluated early 5th instar larvae recorded the maximum yield. The inoculation dose of 1965.87 POB mm‾2 and the incubation temperature of 25°C registered the highest in vivo virus yield. Also, the POB yield was recorded maximum range when virosed larvae were harvested as cadavers. However, among the isolates tested in this study, CMB isolate collected from Tamil Nadu, India, showed the highest yield in all of the conditions tested compared to the other HaNPV isolates.
REFERENCES
- Bell, M.R., 1991. In vivo production of a nuclear polyhedrosis virus utilizing a tobacco budworm multicellular larval rearing container. J. Entomol. Sci., 26: 69-75.
Direct Link - Cherry, A.J., M.A. Parnell, D. Grywacz and K.A. Jones, 1997. The optimization of vivo nuclear polyhedrosis virus production of Spodoptera exempta and S. exigua. J. Inverteb. Pathol., 70: 50-58.
Direct Link - Dolinski, C. and L.A. Lacey, 2007. Microbial control of arthropod pests of tropical tree fruits. Neotrop. Entomol., 36: 161-179.
Direct Link - Huang, Y., 1995. Studies on the propagation of the nuclear polyhedrosis virus of the cotton leafoworm, Spodoptera litura (F.). Acta Scientiarum Naturalium Universitatile Sunyat Seni, 4: 65-69.
Direct Link - Ignoffo, C.M. and M. Shapiro, 1978. Characteristics of baculovirus preparations processed from living and dead larvae. J. Econ. Entomol., 71: 186-188.
CrossRefDirect Link - Kumar, C.M. and R.J. Rabindra, 2003. Influence of dietary vegetable oils on the tobacco cutworm, Spodoptera litura (Fabricius) and its nuclear polyhedrosis virus production. J. Biol. Control, 17: 57-61.
CrossRefDirect Link - Monobrullah, M.D. and M. Nagata, 2000. Optimization of Spodoptera litura (Fab.) nucleopolyhedrovirus production in homologous host larvae. Insect Sci. Applied, 20: 157-165.
Direct Link - Narayanan, K. and S. Jayaraj, 2002. Mass production of polyhedral occlusion bodies of NPV of Helicoverpa armigera in relation to dose, age and larval weight. Indian J. Exp. Biol., 40: 846-849.
Direct Link - Rabindra, R.J., N. Geetha, D. Grzywacz and M. Brown, 2003. Comparative Efficacy of Two Isolates of Nuclear polyhedrosis virus Against Helicoverpa armigera (Hbn.). In: Microbial Pesticides and Insect Pest Management, Rananavare, H.D., S.R. Naik and T.K. Dongre (Eds.). Gandhinagar, Gujarat, India, pp: 127-131.
- Shorey, H.H. and R.L. Hale, 1965. Mass rearing of the larvae of nine noctuid species on a simple artificial medium. J. Econ. Entomol., 58: 522-524.
CrossRefDirect Link - Subramanian, S., G. Santharam, N. Sathiah, J.S. Kennedy and R.J. Rabindra, 2006. Influence of incubation temperature on productivity and quality of Spodoptera litura nucleopolyhedrovirus. Biol. Control, 37: 367-374.
Direct Link