
Nayer Mohammadkhani
Department of Biology, Faculty of Science, Urmia University, Iran
Reza Heidari
Department of Biology, Faculty of Science, Urmia University, Iran
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 21 | Page No.: 3835-3840







ABSTRACT
Response of antioxidative defense system to drought stress was studied in two cultivars of maize differing in sensitivity to drought. The six-days old seedlings were transferred in to different concentrations of polyethyleneglycol (PEG) 6000 as drought stress treatment. After 24 h treatment with PEG 6000 an increase was observed in protective enzymes, MDA content and electrolyte leakage. Under drought stress the activities of protective enzymes in roots and shoots increased sharply. This increase in 704 variety was higher than 301 variety. The protective enzyme activities in roots were lower than those in shoots. The content of malondialdehyde (MDA) increased according to severity of water stress. The content of MDA in roots was lower than in shoots. The electrolyte leakage increased greatly in drought stress, too. Increase of MDA content and electrolyte leakage in 704 variety were higher than 301 variety.
PDF Abstract XML References Citation
How to cite this article
Nayer Mohammadkhani and Reza Heidari, 2007. Effects of Drought Stress on Protective Enzyme Activities and Lipid Peroxidation in Two Maize Cultivars. Pakistan Journal of Biological Sciences, 10: 3835-3840.
DOI: 10.3923/pjbs.2007.3835.3840
URL: https://scialert.net/abstract/?doi=pjbs.2007.3835.3840
DOI: 10.3923/pjbs.2007.3835.3840
URL: https://scialert.net/abstract/?doi=pjbs.2007.3835.3840
INTRODUCTION
Maize (Zea mays L.) is a main food and economical crop. It is one of the most important crops throughout the world. So, it is urgent to improve maize yields even under the unfavorable conditions. Depending on different water stress conditions, biochemical changes to various extent, occurs in maize. Maize production suffers during severe drought (Ti-da et al., 2006).
Drought stress is one of the important limiting factors in plant growth which has limited the production in 25% of arable lands in the world (Levitt, 1980).
Water deficiency can induce membrane damage, increase membrane permeability and the accumulation of free radicals in plants. As a reaction to this condition, antioxidative enzymes can be produced to remove those active oxygen radicals (Sun et al., 2003; Wang et al., 2002).
Higher plants have active oxygen-scavenging systems consisting of several antioxidant enzymes, such as ascorbate peroxidase (APX), guaiacol peroxidase (GPX) and catalase (CAT). These systems protect membranes from the deleterious effects of ROS, such as superoxide radicals, hydrogen proxide (H2O2), hydroxyl radicals and singlet oxygen, which are produced at elevated rates when plants are exposed to abiotic stress conditions (Bowler et al., 1992; Noctor and Foyer, 1998). The superoxide radical is dismutated to H2O2 by SOD, CAT and APX metabolize H2O2 into H2O.
Numerous studies have indicated that activities of antioxidant enzymes are correlated with plant tolerance to abiotic stresses. The drought-induced damage was negatively correlated with the capacity for increase of SOD and CAT activities in mosses cultivars differing in drought tolerance (Dhindsa and Matowe, 1981).
A correlation between antioxidant enzyme activity and water stress or salinity tolerance was demonstrated by comparison of a tolerant cultivar with a sensitive cultivar in several plant species, such as rice (Dionisio-Sense and Tobita, 1998; Srivalli et al., 2003; Vaidyanathan et al., 2003) and tomato (Mittova et al., 2002).
MDA is a major cytotoxic product of lipid peroxidation and has been used widely as an indicator of free radical production (Hong et al., 2000; Hua et al., 1997; Kavikishor et al., 1995; Kunert and Ederer, 1985; Peng et al., 1996).
Electrolyte leakage reflects the damage of stresses to the plasmalemma.
The response mechanisms of reactive oxygen scavering system and lipid peroxidation to drought stress are very complex, as they are not only dependent on plant genotype and stress intensity, but also there are different modes of enzyme reaction.
The main aim of this study was to understand the responses and adaptations of maize to drought and the function of some physiological and biochemical processes.
MATERIALS AND METHODS
Plant materials and growth conditions: This study was conducted at biochemistry laboratory, Department of Biology, Urmia University, Iran, during the winter of 2007.
Two genotypes of corn (Zea mays L.) were used: var. 704 and var. 301 which was obtained from the Agricultural Research Center of Ardebil, Iran.
Seeds were germinated in Petri dishes on two layers of filter paper for three days at 25°C in an incubator. After three days, the seedlings were sown in plastic pots (15 cm diameter, 20 cm depth) filled with sand and irrigated with half strength of Hoagland nutrient solution (Hoagland and Arnon, 1938).
Three uprooted six-days old seedlings transferred to hydroponic culture of aerated test tubes containing polyethyleneglycol (PEG 6000) solutions of 10, 20, 30 and 40% strengths to achieve water deficit levels of -0.15, -0.49, -1.03 and -1.76 Mpa, respectively (Burlyn et al., 1973; Nicholas, 1989; Steuter, 1981) as treatments and aerated test tubes containing half strength Hoagland nutrient solution which served as control. Test tubes were placed in greenhouse for 24 h and thereafter MDA content, electrolyte leakage and protective enzymes activity was determined in roots and shoots.
Measurement of MDA content: MDA content was measured using a 2-thiobarbituric acid reaction (Heath and Packer, 1968; Hong et al., 2000; Chen et al., 2004). Thus 1 g of fresh tissue was homogenized in 5 mL of 5% (W/V) trichloroacetic acid and the homogenate was centrifuged at 10000 g for 15 min at room temperature. The supernatant was mixed with an equal volume of 2-thiobarbituric acid (0.5% in 20% [w/v] trichloroacetic acid) and the mixture was boiled for 25 min at 95°C, followed by centrifugation for 5 min at 7500 g to clarify the solution.
Absorbance of the supernatant was measured at 532 nm and corrected for non-specific turbidity by subtracting the A600. MDA contents were calculated using and extinction coefficient of 155 m-1cm-1.
Determination of electrolyte leakage: The electrolyte leakage was determined as described before (Huang et al., 2005). Shoots and roots of four seedlings were immersed in 15 mL of distilled water in a test tube overnight at room temperature. The initial conductivity was determined using a conductivity meter (model kent EIL5007). The test tubes were then placed in boiling water for 15 min and cooled to room temperature. Conductivity was again determined. The electrolyte leakage was calculated as the ratio of conductivity before boiling to that after boiling.
Determination of enzyme activity: For extraction of plant extracts, 0.5 g fresh weight of shoots and roots were ground in a mortar with pestle in 5 mL buffer (0.05 M tris-HCl buffer, pH = 7.5, 3 mM MgCl2, 1 mM EDTA). Enzyme extracts used for determination of ascorbate peroxidase (APX) activity involve 2 mM ascorbate, too. The homogenate was centrifuged at 5000 g for 20 min. The supernatant was used for determination of GPX, APX and CAT activities (Garratt et al., 2002).
The reaction solution of GPX contained 50 mM phosphate buffer (pH = 7), 1 mL guaiacol 1% and 0.3 mL enzyme extracts. After equilibration at 30°C for 1 min, the reaction was started by the addition of 1 mL H2O2 1% (w/v). GPX activity was calculated by following the decrease in absorbance of tetraguaiacol (extinction coefficient 26.6 mM-1cm-1) within 1 min at 420 nm by UV-visible spectrophotometer (WPA model S2100) (Upadhaya et al., 1985).
The 3 mL reaction solution of APX contained 50 mM phosphate buffer (pH = 7), 0.2 mL H2O2 1% (w/v), 0.5 mM ascorbate, 0.1 mM EDTA and 0.1 mL enzyme extracts. APX activity was calculated by following the decrease in absorbance of ascorbate (extinction coefficient 2.8 mM-1 cm-1) within 1 min at 240 nm by UV -visible spectrophotometer (WPA model S2100) (Asada and Chen, 1989).
The 3 mL reaction solution of CAT contained 50 mM phosphate buffer (pH = 7), 0.2 mL H2O2 1% (w/v) and 50 μL of enzyme extracts. The reaction was initiated by adding enzyme extracts. The decrease of absorbance of H2O2 (extinction coefficient 0.036 mM-1 cm-1) within 1 min at 240 nm was recorded by UV-visible spectrophotometer (WPA model S2100). One unit of CAT activity was defined as the amount of enzyme required to oxidize 1 μM of H2O2 per minute (Maehly and Chance, 1959).
Statistical analyses: Mean values were calculated from measurements of four replicates and the SE of the means were determined. One-way ANOVA and Tukey HSD s multiple range test (p<0.05) was applied to determine the significance of the result between different treatments. All statistical analyses were done using the Statistical Package for Social Sciences (SPSS) for Windows (version 13.0.0).
RESULTS
Effect of water stress on lipid peroxidation (MDA content): MDA is one of the end products which is produced as a result of lipid peroxidation damage by free radicals.
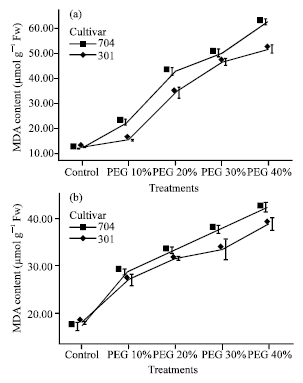 | |
| Fig. 1: | Effects of different PEG 6000 treatments on lipid peroxidation (MDA content) in roots (a) and shoots (b) of two maize cultivars. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
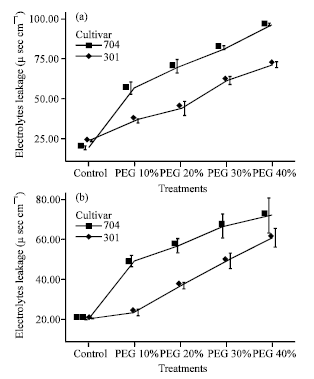 | |
| Fig. 2: | Effects of different PEG 6000 treatments on electrolyte leakage in roots (a) and shoots (b) of two maize cultivars Results are shown as mean±standard error (p<0.05), obtained from four replicates |
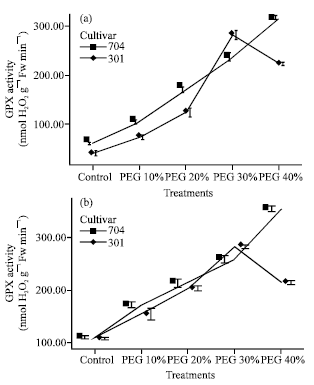 | |
| Fig. 3: | Effects of different PEG 6000 treatments on guaiacol peroxidase (GPX) activity in roots (a) and shoots (b) of two maize cultivars Results are shown as mean±standard error (p<0.05), obtained from four replicates |
Under drought stress the MDA content in roots and shoots both increased depending on drought intensity and plant organ.
Increase of MDA content in 704 var. was higher than in 301 var. and in roots it was higher than in shoots because of high drought stress (Fig. 1).
Increases of MDA content in roots in PEG 10% and in shoots in PEG 20% were enormous.
Effect of water stress on electrolyte leakage: Electrolyte leakage reflected the damage of stresses to the plasmalemma. It showed minor variations in var. 301. Electrolyte leakage in var. 704 increased greatly, which indicated that 301 plants were less damaged under drought stress than 704 plants (Fig. 2).
Electrolyte leakage in roots was increased greater than in shoots. Therefore, the damage of plasmalemma in roots was greater than in shoots, because drought stress in roots was higher than in shoots.
Effects of water stress on protective enzymes activities:
Under water stress, protective enzymes (GPX, APX and CAT) in maize roots and shoots increased in both varieties.
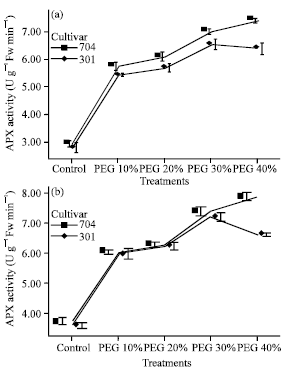 | |
| Fig. 4: | Effects of different PEG 6000 treatments on ascorbate peroxidase (APX) activity in roots (a) and shoots (b) of two maize cultivars Results are shown as mean±standard error (p<0.05), obtained from four replicates |
Under PEG treatments, the activities of APX and CAT in shoots and roots initially increased drastically. This increase was obvious at PEG 10%. In PEG 10% the activity of APX in roots increased 1.8 fold for 704 var. and 1.6 fold for 301 var. as compared to the control and in shoots increased 1.5 fold as compared to the control in both varieties (Fig. 4).
In PEG 10%, the activity of CAT in roots increased 2.6 fold in both varieties and in shoots increased 1.8 fold for 704 var. and 1.5 fold for 301 var. (Fig. 5).
The activity of GPX increased gradually in different treatments (Fig. 3).
In 301 var., protective enzyme activity decreased in PEG 40%. Decrease in APX activity was lower than GPX and CAT. Therefore, 301 plants at PEG 30% showed greater protective enzyme activity than others, but 704 plants at PEG 40% showed high enzymes activity.
In 704 plants enzyme activity and their increase was higher than 301 plants, but in 301 plants APX activity in roots and shoots at PEG 30% was higher than 704 plants.
The protective enzymes activities in roots were lower than those in shoots, but increase in enzyme activity in roots were higher than in shoots. Because, drought was higher in the roots than in the shoots.
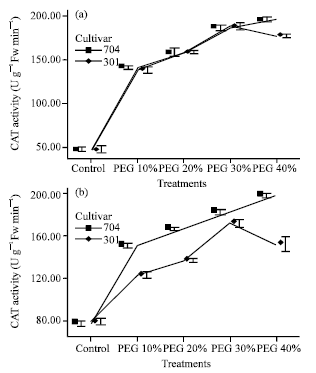 | |
| Fig. 5: | Effects of different PWG 6000 treatments on catalase (CAT) activity in roots (a) and shoots (b) of two maize cultivators. Results are shown as mean±standard error (p<0.05), obtained from four replicates |
DISCUSSION
Numerous investigations have demonstrated that the cellular injury to plants by abiotic stresses is oxidative damage (Asada, 1999; Bowler et al., 1992).
Active oxygen was accumulated in plants under water stress. There is a defensive system in plants, that is to say, plants have an internal protective enzyme- catalyzed cleanup system, which is fine and elaborate enough to avoid injuries of active oxygen, thus guaranteeing normal cellular function (Wang et al., 2002).
When the plants suffered from water stress, the whole defensive system needs to be activated in order to resist the active oxygen injury (Ti-da et al., 2006). Protective enzymes and antioxidants have important roles in protection of plants from oxidative damage.
In the present study was aimed to investigate the responses of Zea mays L. var. 704 and var. 301 to drought stress.
Under PEG treatments, GPX, APX and CAT activities of 704 plants in roots and shoots were not only higher than the control, but were also higher than their activities in 301. Only 301 plants GPX activity in PEG 30% were higher than 704 plants and highest in 301 plants. We have a decrease in enzyme activity of 301 plants in PEG 40%.
There was a linear and significant increase in GPX, APX and CAT activities in both varieties in response to increased water stress, only we have a decrease in PEG 40% in 301 plants. Therefore, there were positive and strong correlation between antioxidant enzymes and drought stress.
Protective enzyme activities in roots were lower and also more sensitive than those in shoots. Drought probably acted directly on roots, whereas leaves could reduce water transpiration by curling and stoma regulation.
In 704 plants MDA production in roots and shoots increased higher than 301 plants. Lower MDA induction was observed in 301 var. suggesting a better protection from oxidative damage as compared with 704 var. The higher level of protection in 301 var seems to be a result of the more efficient antioxidative system, while significant increases in MDA levels in roots and shoots of 704 var. appeared to be derived from low level of protection.
The increased electrolyte leakage was positively correlated with accumulation of MDA, indicating that the plasmalemma injury caused by drought stress is resulted from oxidative damage. In 704 plants, increase in electrolyte leakage in roots and shoots is higher than 301 plants. Therefore in water stress 704 plants were damaged higher than 301 plants and this damage in PEG 40% (-1.76 MPa) is higher than others.
Based on the present results, it appears that plants of the 301 variety have a better tolerance to water stress as compared to 704 variety.
CONCLUSIONS
The results clearly suggest that in maize, under drought stress, in different concentrations of PEG, the activities of GPX, APX and CAT in roots and shoots were increased. Membrane lipid peroxidation and electrolyte leakage increased, too. This increase in 704 plants is higher than 301 plants.
Direct injury to cell membrane system maybe indicating one of the physiological response mechanisms to drought condition. The change of activities of the protective enzymes and MDA and electrolyte leakage are different at different PEG concentrations because of intensity of drought stress.
The increase of protective enzymes activities in roots was higher than shoots in both varieties. Increase of MDA content and particularly increase of electrolyte leakage in roots were higher than shoots in both varieties. These show that intensity of drought stress in roots was higher than shoots and the roots were damaged greater than shoots in both varieties.
ACKNOWLEDGMENTS
This work was supported by the Department of Biology of Urmia University, Iran. We greatly acknowledge Dr. Ilkhanipoor for her support and help in providing materials. We greatly acknowledge the help in modifying this paper from Prof. Carapetian.
REFERENCES
- Chen, G.X. and K. Asada, 1989. Ascorbate peroxidase in tea leaves: Occurrence of two isozymes and the differences in their enzymatic and molecular properties. Plant Cell Physiol., 30: 987-998.
Direct Link - Asada, K., 1999. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol., 50: 601-639.
CrossRefPubMedDirect Link - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Michel, B.E. and M.R. Kaufmann, 1973. The osmotic potential of polyethylene glycol 6000. Plant Physiol., 51: 914-916.
CrossRefPubMedDirect Link - Chen, C.T., T.H. Chen, K.F. Lo and C.Y. Chiu, 2004. Effects of proline on copper transport in rice seedlings under excess copper stress. Plant Sci., 166: 103-111.
Direct Link - Dionisio-Sese, M.L. and S. Tobita, 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci., 135: 1-9.
CrossRefDirect Link - Garratt, L.C., B.S. Janagoudar, K.C. Lowe, P. Anthony, J.B. Power and M.R. Davey, 2002. Salinity tolerance and antioxidant status in cotton culture. Free Radic. Biol. Med., 33: 502-511.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Hong, Z., K. Lakkineni, Z. Zhang and D.P.S. Verma, 2000. Removal of feedback inhibition of Δ1-pyrroline-5-carboxylate synthetase results in increased proline accumulation and protection of plants from osmotic stress. Plant Physiol., 122: 1129-1136.
Direct Link - Huang, M. and Z. Guo, 2005. Responses of antioxidative system to chilling stress in two rice cultivars differing in sensitivity. Biologia Planta, 49: 81-84.
Direct Link - Kishor, P.B.K., Z. Hong, G.H. Miao, C.A.A. Hu and D.P.S. Verma, 1995. Overexpression of δ-pyrroline-5-carboxylate synthetase increases proline production and confers osmotolerance in transgenic plants. Plant Physiol., 108: 1387-1394.
Direct Link - Kunert, K.J. and M. Ederer, 1985. Leaf aging and lipid peroxidation: The role of antioxidants vitamin C and E. Physiol. Plant, 65: 85-88.
CrossRef - Mittova, V., M. Tal, M. Volokita and M. Guy, 2002. Salt stress induces up-regulation of an efficient chloroplasts antioxidant system in the salt-tolerant wild tomato species Lycopersicon pennellii but not in the cultivated species. Plant Physiol., 115: 393-400.
Direct Link - Money, N.P., 1989. Osmotic pressure of aqueous polyethylene glycols: Relationship between molecular weight and vapor pressure deficit. Plant Physiol., 91: 766-769.
CrossRefDirect Link - Noctor, G. and C.H. Foyer, 1998. Ascorbat and glutathione: Keeping active oxygen under control. Ann. Rev. Plant Physiol. Plant Mol. Biol., 49: 249-279.
PubMedDirect Link - Peng, Z., Q. Lu and D.P.S. Verma, 1996. Reciprocal regulation of delta 1-pyrroline-5-carboxylate synthetase and proline dehydrogenase genes controls proline levels during and after osmotic stress in plants. Mol. Gen. Genet., 253: 334-341.
PubMedDirect Link - Srivalli, B., G. Sharma and R. Khanna-Chopra, 2003. Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Plant Physiol., 119: 503-512.
Direct Link - Steuter, A.A., A. Mozafar and J. Goodin, 1981. Water potential of aqueous polyethylene glycol. Plant Physiol., 67: 64-67.
Direct Link - Ge, T.D., F.G. Sui, L.P. Bai, Y.Y. Lu and G.S. Zhou, 2006. Effects of water stress on the protective enzymes and lipid peroxidation in roots and leaves of summer maize. Agric. Sci. China, 5: 291-298.
CrossRefDirect Link - Upadhyaya, A., D. Sankhla, T.D. Davies, N. Sankhla and B.N. Smith, 1985. Effect of paclobutrazol on the activities of some enzymes of activated oxygen metabolism and lipid peroxidation in senescing soybean leaves. J. Plant Physiol., 121: 453-461.
CrossRefDirect Link - Vaidyanathan, H., P. Sivakumar, R. Chakrabarty and G. Thomas, 2003. Scavenging of reactive oxygen species in NaCl-stressed rice (Oryza sativa L.)-differential response in salt-tolerant and sensitive varieties. Plant Sci., 165: 1411-1418.
CrossRefDirect Link