
Ertugrul Aksekili
Department of Biology, Faculty of Arts and Sciences, University of Ondokuz Mayrs,
55139, Kurupelit-Samsun, Turkey
Dudu Duygu Kilic
Department of Biology, Faculty of Arts and Sciences, University of Ondokuz Mayrs,
55139, Kurupelit-Samsun, Turkey
Hamdi Guray Kutbay
Department of Biology, Faculty of Arts and Sciences, University of Ondokuz Mayrs,
55139, Kurupelit-Samsun, Turkey
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 21 | Page No.: 3778-3785







ABSTRACT
Foliar mass per area (mg dm-2) -based nitrogen and phosphorus concentrations, specific leaf mass (mg dm-2) and absolute and proportional resorption in Quercus brantii was investigated along a topographic gradient from 450 to 850 m altitude. Foliar N and P concentrations in Q. brantii exhibited significant differences with respect to topographic position and sampling dates in all of the studied parameters. A sharp decrease was observed from April to September in terms of Specific Leaf Mass (SLM) values. However, mass per area-based absolute and proportional N resorption was lowest at 450 m, while absolute and proportional P resorption was lowest at 850 m. Significant correlations were found between mass per area-based leaf nutrient concentration and foliar resorption except for the correlations between absolute P resorption and foliar P concentrations at 450, 650 and 750 m during full-leaf expansion and 850 m during senescence, respectively.
PDF Abstract XML References Citation
How to cite this article
Ertugrul Aksekili, Dudu Duygu Kilic and Hamdi Guray Kutbay, 2007. Foliar Nutrient Dynamics and Foliar Resorption in Quercus brantii Lindley along an Elevational Gradient. Pakistan Journal of Biological Sciences, 10: 3778-3785.
DOI: 10.3923/pjbs.2007.3778.3785
URL: https://scialert.net/abstract/?doi=pjbs.2007.3778.3785
DOI: 10.3923/pjbs.2007.3778.3785
URL: https://scialert.net/abstract/?doi=pjbs.2007.3778.3785
INTRODUCTION
The resorption of nutrients prior to leaf fall is one of the key processes by which plants conserve them. This process reduces the likelihood of nutrient loss in litter dropped on the forest flor and subsequently, the withdrawn nutrients are redeployed in new tissues, such as leaves and reproductive structures or stored for later use. The resorption accompanied by a reduction in nutrient restitution (through leaf litter) and requirements, affords the ecosystem a certain independence from the soil and the possibility of good management of the available elements (Singh et al., 2005).
Forest trees and shrubs retranslocate sizeable portions of the nutrient contents of leaves before leaf senescence. One of the most important methods to measure nutrient use efficiency in plants is to determine foliar resorption, the process of nutrient translocation from the leaves into storage tissues during senescence (Killingbeck, 1986; Killingbeck and Costigan, 1988). Nutrient resorption reduces nutrient loss with annual litterfall, so that it reduces dependence on soil nutrient supply (Xue and Luo, 2002).
Both endogenous and exogenous factors, cause foliar nutrients to vary in time and space (Del Arco et al., 1991). The main cause of spatial variation is topography, vegetation, structure and disturbance. Temporal variation is primarily regulated by leaf age and growth, overall plant phenological state and weather (Chabot and Hicks, 1982; Oleksyn et al., 2002).
Grime (2002) suggested that mineral nutrients, especially nitrogen and phosphorus should be considered as the major regulating factors of vegetation processes at scales ranging from the individuals to ecosystems. Oleksyn et al. (2002) concluded that intraspecific genetic differences exist in foliage nutrient concentrations among diverse populations and if nutrient availability were limited as a result of interactions between temperature, litter quality and mineralization, a tendency toward higher foliage concentrations of macronutrients can be adaptive feature enhancing plants metabolic activity in their native habitats.
In deciduous species, foliar nutrient concentrations will remain relatively constant from the time of full leaf expansion to the beginning of senescence, then decrease rapidly as nitrogen and phosphorus are resorbed prior to abscission (Hevia et al., 1999).
Foliar nutrient content is widely recognized as an effective measure of the nutritional status of plants because leaves are the primary sites of physiological activity including photosynthesis, respiration, transpiration, gas exchange and nutrient storage (Oleksyn et al., 2002).
Quercus L. comprises 531 tree and shrub species distributed among contrasting phytoclimates in the Northern Hemisphere, from temperate and subtropical deciduous forests to Mediterranean evergreen woodlands (Corcuera et al., 2002). Quercus brantii Lindley is a deciduous shrub or small tree up to a height of 6-10 m belonging to Fagaceae and distributed in Irano-Turanien phytogeographic region (Davis, 1982). We selected this species for our investigation because it occurs along the entire topographic gradient and it is a typical species of the Irano-Turanien phytogeographic region forming pure stands.
The main aim of this study is to examine whether mass per leaf area (mg dm-2) -based nitrogen and phosphorus concentrations, specific leaf mass (mg dm-2) and absolute and proportional resorption changed the topographic position.
MATERIALS AND METHODS
The study area and the collection of samples: Q. brantii specimens were collected from Batman city in the southeastern part of Turkey (41°29’ E; 38°23’N) from plots located at altitudes from 450 to 850 m. The plots are located at the B8 square according to the grid system of Davis (1982). Q. brantii formed pure stands in the study area. Mean annual temperature in the study area is 15.8°C and mean annual precipitation 552.2 mm. Maximum temperature in the hottest month; July (M) is 44.1°C and minimum temperature for the coldest month; January (m) -19.4°C. Pluviometric quotient (Q) is 33.43. Mean annual relative humidity in summer and winter are 80 and 31%, respectively. According to these data the study area has a continental climate (Anonymous, 1994).
Plots of 20x20 m were sampled along a topographic gradient altitudes from 450 to 850 m (450, 550, 650, 750 and 850 m) above sea level. Plots were selected to have stands with closed tree canopies on southwest facing slopes. From each topographic position five Q. brantii individuals (≥4.5 m tall) were randomly selected and flagged. Individuals were selected a ≥3 m distance from the stems of neighboring canopy trees to avoid potential microsite variation (Boerner and Koslowsky, 1989). Since sun and shade leaves may differ in foliar nutrient concentrations, only sun leaves were collected (Grubb, 1977).
Deciduous forest species usually reach full-leaf expansion phase during earlysummer to midsummer (Diaz and Cabido, 1997). Mature leaves were preselected and they are to be collected when they have senesced. Preselection also minimises the chance of comparing green and senesced leaves of different cohorts (Van Heerwarden et al., 2003). Five fresh leaves from throughout the midcrown per individual were collected to avoid effects of crown position at the beginning of leaf expansion (at the second half of April 2000) and at the full-leaf expansion (mid-growing season) leaves (at the first half of July 2000). Senesced leaves (five leaves) were also collected at the second half of September 2000. Leaf area of specimens was measured with a leaf area meter (LI-3000, LI-COR, USA). Leaf dry mass was determined after oven-drying at 70°C for at least 3 days. After that specific leaf mass (SLM as mg dm-2) was calculated as the leaf dry mass per unit leaf area.
From each topographic position five soil samples were taken at a depth of 30 cm with a soil corer. Each sample from each topographic position was a composite of five subsamples collected at random beneath the plant specimens. Soil samples were air-dried and sieved to pass through a 2 mm mesh prior to analysis. Because soil parameters changed very little during both sampling period soil samples belonging to both sampling period were pooled.
Methods of chemical analysis: Leaf samples were dried at 70°C until constant weight, ground, sieved and digested in a mixture of nitric and perchloric acids with the exception of samples for N analysis. Nitrogen (N) and phosphorus (P) analysis were made by standard methods (Allen et al., 1986).
Foliage nutrient concentrations were multiplied by leaf mass and absolute resorption was calculated as the difference between the concentrationxleaf mass of the nutrient in fresh and senesced-leaves (Staaf, 1982; Wright and Westoby, 2003). As leaves can lose as much as 30 % their total dry weight during senescence and resorption estimates based on the quantity of nutrient per unit leaf area are more accurate than those determined on a mass basis. Proportional resorption was then calculated as the percentage reduction in leaf N and P per unit leaf area from fresh to senesced-leaves (Lusk and Contreras, 1989; Wright and Westoby, 2003).
Soil pH, soil nitrogen (g kg-1) and soil phosphorus (g kg-1) were determined by standard methods. For the determination of soil moisture (cm3 H2O/100 cm3 soil) about 250-300 g samples were placed in soil pins, weighed fresh, dried at 105°C for 48 h; then weighed dry and then soil moisture was calculated on a volume basis (Boerner, 1984). The results of soil analysis were explained according to Bayraklı (1987).
Statistical analyses: One-way ANOVA test was performed to determine whether or not SLM (mg dm-2) change with respect to leaf growth stage by using SPSS 10.0 version (Anonymous, 1999; Schaefer and Anderson 1989). Two-way ANOVA test was also performed to determine whether or not N, P and SLM change in terms of leaf growth stage and elevation by using MINITAB 12.1 version (Anonymous, 1998). Dependent and independent variables were N and P concentrations and SLM and leaf growth stages and elevation, respectively.
Multiple comparisons among pairs of means were performed using Duncan’s multiple range test when a significant ANOVA result occurred.
| Table 1: | Variance analysis in terms of mass (mg g-2) and mass per area (mg dm-2)-based N and P concentrations and SLM (mg dm-2) of Q. brantii leaves with respect to elevation and months by two-way ANOVA |
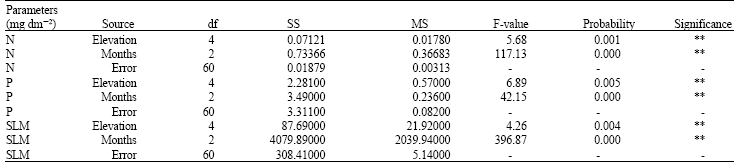 | |
| df:: Degrees of freedom, SS: Sum of Squared, MS: Mean Square, *: p<0.05 **: p<0.01 | |
RESULTS
Foliar N and P concentrations in Q. brantii exhibited significant differences with respect to topographic position and sampling dates in all of the studied parameters (Table 1). Mass per leaf area-based N concentrations were higher at 850 m while the lowest mass per leaf area-based N concentrations were found at 450 m. The highest and the lowest mass per leaf area-based P concentrations were found at 550 and 850 m, respectively. In summary, Mass per leaf area-based N concentrations were increased with altitude, while mass per leaf area-based P concentrations were decreased with altitude. The highest and the lowest SLM values were observed at 550 and 850 m, respectively like mass per leaf area-based P concentrations (Fig. 1-4). SLM values were slightly increased from April to July. However, a sharp decrease was observed during September. According to the results of Duncan’s multiple range test upland and bottomland positions were significantly different from each other in terms of N and P (mg dm-2) concentrations and SLM (mg dm-2) (Table 2).
Leaves showed a maximum N and P concentrations and SLM in April and nutrient concentrations and SLM rapidly dropped from April to June (Table 3).
Significant differences were observed along the topographic gradient with respect to absolute and proportional foliar resorption. According to the results of Duncan’s multiple range test upland positions (850 m) were significantly different from bottomland positions (450 m). Mass per leaf area-based proportional N resorption was lowest at 450 m, while proportional P resorption was lowest at 850 m (Table 4).
Significant correlations were found between mass per leaf area-based leaf nutrient concentration and foliar resorption. However, the correlations between mass per leaf area-based absolute P resorption and foliar P concentration at 550 and 650 m during full-leaf expansion and 750 and 850 m during senescence were not statistically significant (Table 5 and 6).
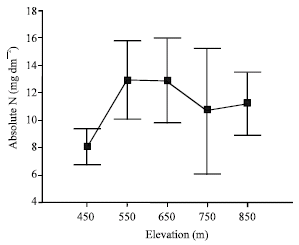 | |
| Fig. 1: | Mass per leaf area-based absolute N (mg dm-2) resorption along the topographic gradient |
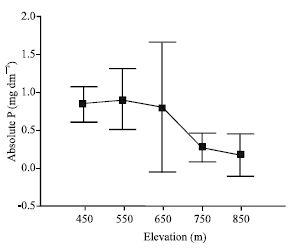 | |
| Fig. 2: | Mas per leaf area-based absolute P (mg dm-2) resorption along the topographic gradient |
Soil N and P concentrations were low to medium levels at 450, 550 and 850 m although soil P and N concentrations were higher at slope positions (650 and 750 m, respectively). Similarly soil moisture were higher at slope positions (650 and 750 m). There were significant differences in all of the soil parameters along the topographic position except for soil pH (Table 7).
| Table 2: | The comparison of mass per leaf area (mg dm-2) based N and P concentrations and SLM (mg dm-2) of Q. brantii leaves on the basis of elevations |
 | |
| Means followed by the same letter(s) are not significantly different at the 0.05 level using Duncan’s multiple range test | |
| Table 3: | The comparison of mass per leaf area (mg dm-2) based N and P concentrations and SLM (mg dm-2) of Q. brantii leaves on the basis of months |
 | |
| Means followed by the same letter(s) are not significantly different at the p<0.05 level using Duncan’s multiple range test | |
| Table 4: | Mass per leaf area-based absolute and proportional N and P resorption along topographic gradient in Q. brantii. Standard errors were given in parenthesis |
 | |
| Means followed by the same letter(s) are not significantly different at the 0.05 level using Duncan’s multiple range test. *: p<0.05 **:p<0.01 | |
| Table 5: | Pearson correlation coefficients between mass per leaf area-based absolute and proportional resorption and foliar N and P concentrations during full-leaf expansion |
 | |
| **: p<0.01, NS: Not Significant | |
| Table 6: | Pearson correlation coefficients between mass per leaf area-based absolute and proportional resorption and foliar N and P concentrations during senescence |
 | |
| **: p<0.01, NS: Not Significant | |
| Table 7: | Mean values for soil parameters along the topographic position and the comparison of soil parameters by one-way ANOVA test. Standard errors are given in parenthesis |
 | |
| Means followed by the same letter are not significantly different at the 0.05 level using Tukey’s HSD test. *: p<0.05 NS: Not Significant Sig.: Significance | |
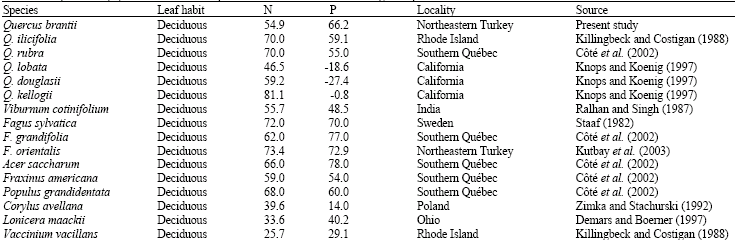
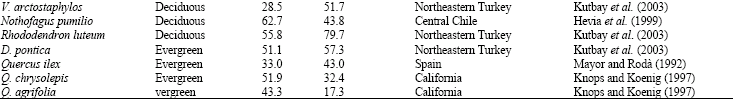
| Table 8: | Proportional (%) N and P foliar resorption for some deciduous and evergreen species |
 | |
 | |
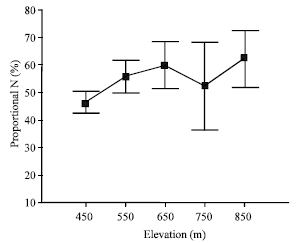 | |
| Fig. 3: | Mass per leaf area-based proportional N (%) resorption along the topographic gradien |
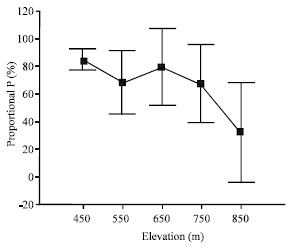 | |
| Fig. 4: | Mass per leaf area-based proportional P (%) resorption along the topographic gradient |
There were some differences between proportional resorption values of Q. brantii and the other deciduous species (Table 8).
DISCUSSION
The first sampling date (at the beginning of leaf expansion) was significantly different from the other sampling dates with respect to foliar N and P concentrations. Nitrogen and phosphorus in particular are highly influenced by seasonal variations (Escudero et al., 1992). Similar differences were also observed in some other deciduous species, e.g., canopy and subcanopy species in a Fagus orientalis Lipsky forest and a Quercus cerris L. var. cerris forest (Kutbay and Kılınc, 1994; Kutbay et al., 2003).
Temporal variation which primarily regulated by leaf age and growth is found to be the most important factor on foliar nutrient contents during biological cycles of leaves (Chabot and Hicks, 1982; Mayor and Rodà, 1992; Orgeas et al., 2002). The vegetative cycle of deciduous forest species undergoes three stages of development: the first is of rapid growth, the second refers to maturation and the third involves senescence. During full-leaf expansion nutrient concentrations increase rapidly and decrease through senescence (Regına and Tarazona, 2001).
Orgeas et al. (2002) stated leaf ageing was more important during the early stages of leaf development, as it was a result of nutrient dilution with leaf growth whereby the rate of nutrient input was less than rapid leaf growth. They also found temporal variation which is regulated by leaf ageing processes was much more important in mineral nutrition than spatial variation which is regulated mainly by disturbance. Leaf senescence is a complex, highly regulated developmental phase in the life of a leaf that results in the coordinated degradation of macromolecules. Nitrogen resorption is the physiological process by which plants withdraw nitrogen from senescing leaves, making it available for investment in new structures (Cartaxana and Catarino, 2002). These findings are also supported by Boerner (1984), Boerner (1985), Eckstein et al. (1999), Feller and Fischer (1994) and Minoletti and Boerner (1994).
Mass per leaf area-based N concentrations were increased with altitude, while mass per leaf area-based P concentrations were decreased with altitude. Tanner et al. (1998) found conflict results in terms of N and P concentrations of tropical montane trees. However, P concentrations of tropical montane trees were decreased with altitude.
SLM is a good indicator of relative adaptability to shade (Bigelow, 1993; Jurik, 1986; Kutbay, 2001). SLM values in the present study were similar to other Quercus L. species. Q. brantii may be classified as a shade-intolerant species because SLM values were rather low as in other Quercus species (Abrams and Kubiske, 1990; Lusk and Contreras, 1989).
Demars and Boerner (1997) and Kutbay and Ok (2003) hypothesized foliar resorption probably reflected topographic differences in soil moisture than in soil fertility. However, the present study was not fully support that hypothesis. The highest soil moisture was observed at 650 and 750 m. Although the highest absolute N resorption was found at 650 m, the highest proportional N resorption and absolute and proportional P resorption was not found at 650 and 750 m.
Demars and Boerner (1997) also hypothesized foliar resorption would be greater in the lower fertility sites. Our results were consistent with that hypothesis except for absolute N resorption. Proportional N resorption was higher at 850 m where soil N concentration was lowest. Soil P concentration was lowest at 450 and 550 m and proportional and absolute P resorption was highest at 450 and 550 m, respectively.
Although, Aerts (1996) pointed out that relations between leaf nutrient status and leaf nutrient resorption were very weak, Hevia et al. (1999) showed such a relationship does exist, at least for N. The observation of significant positive and negative correlations between foliar N and P concentrations and absolute and proportional resorption during both full-leaf expansion and senescence indicated there were some relations between leaf nutrient status and leaf nutrient resorption in Q. brantii. Côté et al. (2002) stated the resorption generally increased with a decrease in leaf N and P concentration. The negative correlations between mass per area based foliar N and P concentrations and absolute and proportional N and P resorption were explained in this way. Côté et al. (2002) also suggested resorption increases with leaf nutrient concentrations in some cases and the increase is more pronounced for leaf P. High and positive correlations were usually found between foliar P concentration and proportional P resorption in the present study.
There were some differences between proportional resorption values of Q. brantii and the other deciduous species although N and P resorption values were usually within the ranges reported by Aerts and Chapin (2000) and Schulze et al. (1994). For instance, proportional P resorption was usually higher than the other Quercus species and P resorption in Q. brantii was more efficient than the other Quercus species. P is a very mobile within the plant and most of the phosphorus was resorbed during senescence (Ralhan and Singh, 1987). In addition to this, P resorption was more efficient than N resorption in Q. brantii in agreement with other studies (Kost and Boerner, 1985; Mayor and Rodà, 1992; Xue and Luo, 2002; Kutbay and Ok, 2003). In other words, Q. brantii translocates greater percentage of P out of leaves before senescence Knops and Koenig (1997) stated evergreen species had higher P resorption than the deciduous species on the contrary to the present study. In addition to this, being a deciduous species proportional N resorption value of Q. brantii was similar to evergreen Q. chrysolepis and Q. agrifolia and lower than the other deciduous Quercus species. Future research should be done to more precise explanation of leaf nutrient status and leaf nutrient resorption in deciduous and evergreen species.
REFERENCES
- Abrams, M.D. and M.E. Kubiske, 1990. Leaf structural characteristics of 31 hardwood and conifer tree species in Central Wisconsin: Influence of light regime and shade-tolerance rank. For. Ecol. Manage., 31: 245-253.
CrossRef - Aerts, R., 1996. Nutrient resorption from senescing leaves of perennials: Are there general patterns?. J. Ecol., 84: 597-608.
Direct Link - Aerts, R. and F.S. Chapin III, 1999. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. Adv. Ecol. Res., 30: 1-67.
CrossRefDirect Link - Bigelow, S.W., 1993. Leaf nutrients in relation to stature and life form in tropical rain forest. J. Veg. Sci., 4: 401-408.
Direct Link - Boerner, R.E.J., 1984. Foliar nutrient dynamics and nutrient use efficiency of four deciduous tree species in relation to nutrient fertility. J. Applied Ecol., 21: 1029-1040.
Direct Link - Boerner, R.E.J. and S.D. Koslowsky, 1989. Microsite variations in soil chemistry and nitrogen mineralization in a beech-maple forest. Soil Biol. Biochem., 21: 795-801.
CrossRef - Cartaxana, P. and F. Catarino, 2002. Nitrogen resorption from senescing leaves of three salt marsh plant species. Plant Ecol., 159: 95-102.
CrossRefDirect Link - Chabot, B.F. and D.J. Hicks, 1982. The ecology of leaf life spans. Ann. Rev. Ecol. Syst., 13: 229-259.
CrossRefDirect Link - Corcuera, L., J.J. Camarero and E. Gil-Pelegr�n, 2002. Functional groups in Quercus species derived from the analysis of pressure-volume curves. Trees, 16: 465-472.
CrossRefDirect Link - C�te, B., J.W. Fyles and H. Djalilvand, 2002. Increasing N and P resorption efficiency and proficiency in northern deciduous hardwoods with decrease foliar N and P concentrations. Ann. For. Sci., 59: 275-281.
CrossRefDirect Link - Del Arco, J.M., A. Escudero and Garrido, 1991. Effects of site characteristics on nitrogen resorption from senescing leaves. Ecology, 72: 701-708.
Direct Link - Demars, B.G. and R.E.J. Boerner, 1997. Foliar nutrient dynamics and resorption in naturalized Lonicera maackii (Caprifoliaceae) populations in Ohio, USA. Am. J. Bot., 84: 112-117.
Direct Link - D�az, S. and M. Cabido, 1997. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci., 8: 463-474.
CrossRefDirect Link - Eckstein, R.L., P.S. Karlsson and M. Weih, 1999. Leaf life span and nutrient resorption as determinants of plant nutrient conservation in temperate-arctic regions. New Phytol., 143: 177-189.
Direct Link - Feller, U. and A. Fischer, 1994. Nitrogen metabolism in senescing leaves. Crit. Rev. Plant Sci., 13: 241-273.
CrossRef - Grime, J., 2001. Plant Strategies, Vegetation Processes and Ecosystem Properties. 2nd Edn., John Wiley and Sons Ltd., Chichester, Pages: 456.
Direct Link - Grubb, P.J., 1977. Control of forest growth and distribution on wet tropical mountains. Ann. Rev. Ecol. Syst., 8: 38-107.
CrossRefDirect Link - Hevia, F., M.L. Minoletti, K.L.M. Decker and R.E.J. Boerner, 1999. Foliar nitrogen and phosphorus dynamics of three Chilean Nothofagus (Fagaceae) species in relation to leaf lifespan. Am. J. Bot., 86: 447-455.
Direct Link - Jurik, T.W., 1986. Temporal and spatial patterns of specific leaf weight in successional Northern hardwood tree species. Am. J. Bot., 73: 1083-1092.
Direct Link - Killingbeck, K.T., 1986. The terminological jungle revisited: Making a case for use of the term resorption. Oikos, 46: 263-264.
CrossRefDirect Link - Killingbeck, K.T. and S.A. Costigan, 1988. Element resorption in a guild of understory shrub species: Niche differentiation and resorption thresholds. Oikos, 53: 366-374.
Direct Link - Knops, J.M.H. and W.D. Koenig, 1997. Site fertility and leaf nutrients of sympatric evergreen and deciduous species of Quercus in central coastal California. Plant Ecol., 130: 121-131.
CrossRefDirect Link - Kost, J.A. and R.E.J. Boerner, 1985. Foliar nutrient dynamics and nutrient use efficiency in Cornus florida. Oecologia, 66: 602-606.
CrossRefDirect Link - Kutbay, H.G. and M. Kilin�, 1994. Sclerophylly in Quercus cerris L. var. cerris and Phillyrea latifolia L. and edaphic relations of these species. Vegetatio, 113: 93-97.
Direct Link - Kutbay, H.G., 2001. Nutrient content in leaves from different strata of a swamp forest from Northern Turkey. Pol. J. Ecol., 49: 221-230.
Direct Link - Kutbay, H.G. and T. Ok, 2002. Foliar N and P resorption and nutrient levels along an elevational gradient in Juniperus oxycedrus L. subsp. macrocarpa (Sibth. and Sm.) Ball. Ann. For. Sci., 60: 449-454.
CrossRefDirect Link - Kutbay, H.G., E. Yalcin and A. Bilgin, 2003. Foliar N and P resorption and foliar nutrient concentrations in canopy and subcanopy of a Fagus orientalis Lipsky forest. Bel. J. Bot., 136: 35-44.
Direct Link - Lusk, C.H. and O. Contreras, 1989. Foliage area and crown nitrogen turnover in temperate rain forest juvenile trees of differing shade tolerance. J. Ecol., 87: 973-983.
CrossRefDirect Link - Mayor, X. and F. Rod�, 1992. Is primary production in holm oak forests nutrient limited?. Vegetatio, 99: 209-217.
CrossRefDirect Link - Minoletti, M.L. and R.E.J. Boerner, 1994. Drought and site fertility effects on foliar nitrogen and phosphorus dynamics and nutrient resorption by the forest understory shrub Viburnum acerifolium L. Am. Midl. Nat., 131: 109-119.
Direct Link - Oleksyn, J., P.B. Reich, R. Zytkowiak, P. Karolewski and M.G. Tjoelker, 2002. Needle nutrients in geographically diverse Pinus sylvestris L. populations. Ann. For. Sci., 59: 1-18.
CrossRefDirect Link - Orgeas, J., J.M. Ourcival and G. Bonin, 2002. Seasonal and spatial patterns of foliar nutrients in cork oak (Quercus suber L.) growing on siliceous soils in Provence (France). Plant Ecol., 164: 201-211.
Direct Link - Ralhan, P.K. and S.P. Singh, 1987. Dynamics of nutrients and leaf mass in central Himalayan forest trees and shrubs. Ecology, 68: 1974-1983.
Direct Link - Regina, I.S. and T. Tarazona, 2001. Nutrient cycling in a natural beech forest and adjacent planted pine in northern Spain. Forestry, 74: 11-28.
CrossRefDirect Link - Schulze, E.D., F.M. Kelliher, C. Korner, J. Lloyd and R. Leuning, 1994. Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate and plant nitrogen nutrition: A global scaling exercise. Ann. Rev. Ecol. Syst., 25: 629-660.
CrossRefDirect Link - Singh, S.P., K. Bargali, A. Joshi and S. Choudhury, 2005. Nitrogen resorption in leaves of tree and shrub seedlings in response to increasing soil fertility. Curr. Sci., 89: 389-396.
Direct Link - Tanner, E.V.J., P.M. Vitousek and E. Cuevas, 1998. Experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology, 79: 10-22.
Direct Link - Van Heerwaarden, L.M., S. Toet and R. Aerts, 2003. Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: Facts and solutions. Oikos, 101: 664-669.
CrossRefDirect Link - Wright, I.J. and M. Westoby, 2003. Nutrient concentration, resorption and lifespan: Leaf traits of Australian sclerophyll species. Funct. Ecol., 17: 10-19.
Direct Link - Xue, L. and S. Luo, 2002. Seasonal changes in the nutrient concentrations of leaves and leaf litter in a young Cryptomeria japonica stand. Scand. J. For. Res., 17: 495-500.
CrossRefDirect Link