
Jemaa Essemine
Departement des Sciences Biologiques, Laboratoire de Physiologie et Biotechnologie Vegetales Faculte des Sciences de Tunis, 1060, Tunisia
Saida Ammar
Departement des Sciences Biologiques, Laboratoire de Physiologie et Biotechnologie Vegetales Faculte des Sciences de Tunis, 1060, Tunisia
Najoua Jbir
Departement des Sciences Biologiques, Laboratoire de Physiologie et Biotechnologie Vegetales Faculte des Sciences de Tunis, 1060, Tunisia
Sadok Bouzid
Departement des Sciences Biologiques, Laboratoire de Physiologie et Biotechnologie Vegetales Faculte des Sciences de Tunis, 1060, Tunisia
Pakistan Journal of Biological Sciences
Year: 2007 | Volume: 10 | Issue: 21 | Page No.: 3762-3768







ABSTRACT
The aim of this study is determine the effect of different temperature regimes on germination of wheat seeds and early germination events. Germination is very sensitive to environment conditions, particularly the temperature. Physiological and biochemical responses of wheat seed germination during time at various temperatures (5, 15, 25, 35 and 45°C) have shown that optimal temperature (25°C) favorites a good aptitude to germinate, whereas low (5°C) and high temperature (45°C) were extend the delay of germination. Residuary dry matter’s mass of germinating, both Karim and Salambô wheat species, Seeds at 5 and 45°C decreased lightly. This show a less mobilization of reserves to embryo. The activity of totals solubles peroxidases changes with temperature of wheat seed germination. At low (5°C) and high (45°C) temperatures, for both two cultivars, peroxidase activity decreases during germination. With exception, during imbibition (2 h), only 45°C involves a high stimulation of this activity for Karim cultivar. This increase is considered as a biochemical response to high temperature. In control temperature (25°C), we have obtained a light increase in peroxidase activity in comparison with that of dry seeds for both these two wheat species. At the end of this study, we have studied the effect of heat stress on totals proteins content. Nevertheless, both for these two wheat cultivars, at 5°C there is no a markedly change in proteins amount during germination. In control condition (25°C) of germination, there is a slight decrease in this content. Germination under high temperature, for Karim variety, induces a rapid synthesis of Heat Shock Proteins (HSPs) and concomitantly a weak degradation of normal proteins. The opposite phenomenon was observed for Salambô variety (weak synthesis of HSPs and important degradation of normal proteins).
PDF Abstract XML References Citation
How to cite this article
Jemaa Essemine, Saida Ammar, Najoua Jbir and Sadok Bouzid, 2007. Sensitivity of Two Wheat Species’s Seeds (Triticum durum, Variety Karim and Triticum aestivum, Variety Salambô) to Heat Constraint During Germination. Pakistan Journal of Biological Sciences, 10: 3762-3768.
DOI: 10.3923/pjbs.2007.3762.3768
URL: https://scialert.net/abstract/?doi=pjbs.2007.3762.3768
DOI: 10.3923/pjbs.2007.3762.3768
URL: https://scialert.net/abstract/?doi=pjbs.2007.3762.3768
INTRODUCTION
In wheat seeds non-optimal temperatures, allowing germination, mainly affect the germination rate (Hampson and Simpson, 1990). Low temperatures are also responsible for an extended delay of the onset of radicle emergence as well as for a slowing down of seedling growth. As temperature becomes too high, the excess of hydration is associated with high temperature lesions, mostly at the level membrane and enzymatic proteins, impairing the germinative process. The high temperature tolerance in germinating wheat seeds has been well documented by Abernethy et al. (1989) and related to the expression of heat shock induced proteins and their protective function. Therefore, in this study we present further comparative studies on this topic with two different species of wheat (Triticum aestivum. cv. Salambô and Triticum durum. cv. Karim). This study reports the effect of different temperature regimes on germination of wheat seeds and also focuses on early germination events. In addition, our aim is to determine the physiological and biochemical responses of wheat seeds to heat constraint during germination.
MATERIALS AND METHODS
Plant material: Seeds of Karim (Triticum durum) and Salambô (Triticum aestivum) cultivars of wheat were obtained from the National Research Agronomic Institute of Tunisia (INRAT). The first cultivar (cv. Karim) results from crossing: 21563 AA «S» x Fg «s» realized in Mexico and introduced in Tunisia at 1973 (Maamouri et al., 1988). The second (cv. Salambô) was obtained by crossing Pato x CO-INIA realized also in Mexico and introduced early in Tunisia (in year 1970). This study was carried out in the laboratory of Plants Physiology and Biotechnology of Sciences Faculty (University of Tunisia El Manar, Tunisia), at years 2003-2004.
Germination conditions: Wheat seeds were germinated, at different temperature (5, 15, 25, 35 and 45°C), in the dark. Germination was measured at various times, until 34 h and was considered to have taken place when the protrusion of radicles was evident. Batches of twenty five seeds were spread for each treatment on 14 cm Petri dishes with 2 layers of whatman filter paper wetted with 20 mL of deionized distilled water.
Germination kinetic: Germination kinetic was monitored every two hours for 34 h and seeds were counted and removed when they exhibited radicle extension ≥ 2 mm. two germination indices were calculated for each treatment replicate: Total percentage germination and time required to reach 50% germination based on the total number of seeds (Hardegree et al., 2002). Time required to achieve 50% germination was calculated by interpolation from the cumulative germination curve (Covell et al., 1986; Brown and Mayer, 1988). The germination tests were conducted at various temperatures at the beginning of imbibition (0 h).
Reversibility test: Wheat seeds do not germinated at low (5°C) and high (45°C) temperatures were transferred to control temperature (25°C). Seeds do not also germinate after transfers to 25°C were considered as to have an undamaged embryo.
Water uptake: This parameter was determined after desiccation of germinated seeds at 80°C during 3 days. It is calculated as the difference between the mass of fresh weight and dry weight reported to dry weight. Water uptake was exported in mL g-1 DW.
Water uptake = FW-DW/DW |
Soluble POD activity determinations: Germinated seeds were ground (5 seeds) using a chilled mortar and pestle in ice-cold 100 mmol L-1 soduim phosphate buffer pH 7.0 using a ratio of 400 μL buffer/100 mg seed fresh mass. Seed homogenates were centrifuged at 13, 000 xg for 15 min at 4°C and the cleared supernatant was used immediately as the enzyme source. Soluble peroxidase activity was analyzed by following the formation of tetraguaïacol in Beckman DU 640 spectrophotometer. Each reaction mixture (1.01 mL) consisted of 10 μL enzyme extract and 900 μL guaïacol (0.1%) solution containing 100 μL guaïacol in 100 mL soduim phosphate buffer pH 6.0 and 0.5% H2O2 (v/v). POD activity in the extracts was measured as an increase in absorbance at 470 nm min-1 g seed fresh mass-1. The reaction was linear with time and followed for 1 min.
Protein content estimations: Protein concentration was estimated by the Bradford (1976) using the Bio-Rad reagent with bovine serum albumin, BSA, (sigma) as the standard.
Statistics: Statistical data analysis was performed by Analysis of Variance (ANOVA). Student’s tests were carried out to test significance of differences between treatment means using the SPSS for Windows, release 10.0.
RESULTS AND DISCUSSION
Effect of temperature on germination kinetic: Germination curves analysis shows that maximum rate of germination (100%) was registered at 25, 35 and 15°C temperatures, with duration equal 14, 22 and 30 h, respectively. Though, this rate is 68% at 5°C and it is only 38% at 45°C, after 34 h (Fig. 1).
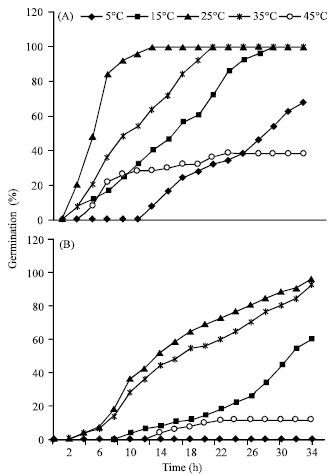 | |
| Fig. 1: | Effect of high and low temperature during germination of intact wheat seed (Karim and Salambô) as shown by cumulative germination over 34 h. The heat shock was administered at the initiation of imbibition (0 h). Values represent the mean of three replicate samples of 25 seeds each. A: Karim; B: Salambô |
| Table 1: | Germination percentage, delay time and germination rate at various temperatures (5-45°C). 1; Triticum durum (var. Karim) 2; Triticum aestivum (var. Salambô) |
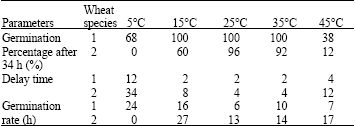 | |
Germination rate for Salambô cultivar is very lower than that of Karim variety. After 34 h, it is 60, 92 and 96% at 15, 35 and 25°C, respectively (Table 1). The rate attains 12% after 22 h of germination at 45°C. On the contrary, of Karim cultivar, no germination was observed for Salambô cultivar after 34 h of germination at 5°C temperature. These results allow that maximum rate of germination was obtained for, enough, a wide range of temperature (15, 25 and 35°C). This is agree with the study of Murphy and Thomas (1982) which showed that radish seeds majority have a thermal optimum comprise between 15 and 30°C.
Results registered in the (Fig. 1) revealed a delay time, for Karim cultivar, in order of two hours at 15, 25 and 35°C temperatures (Table 1). This delay is very important at 5°C (12 h) and at 45°C (4 h). For Salambô cultivar, delay time lasts 4 h at 25 and 35°C. It is 8 and 12 h at 15 and 45°C, respectively. At 5°C, no germination was detected after 34 h. Present result is in agreement with that obtained by (Dell’Aquila and Spada, 1994; Newell and Bludau, 1993). These authors showed that low temperature (5°C) delays radicle protrusion of hard wheat seeds, variety Norba and that imbibition phase extension was related to a process, slow and gradual, of proteins synthesis in wheat embryo.
Reversibility test: Obtained results have shown that, as well as, low (5°C) and high (45°C) temperatures have a depressing effect on wheat seeds germination. A reversibility test should allow exactly if this a consequence of loss viability or to embryo dormancy submitted to an excessive temperature.
Results consigned in Fig. 2 reveale that ulterior seeds transfer from 5°C to control temperature (25°C) favorites a fast germination (100%). That’s why, cold seems to have a benefic effect on the germination and not undamaged the embryo (Mazliak, 1992). On the other hand, seeds transferred from 45-25°C involved retaking germination for only few seeds (45%). Present results confirmed that obtained by Bewley and Black (1982).
Effect of temperature on water uptake during germination: Karim and Salambô dry seeds have a drought content 10 and 7%, respectively (Table 2).
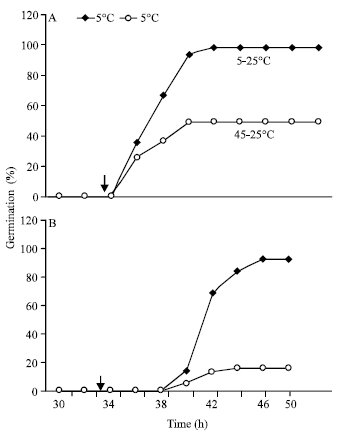 | |
| Fig. 2: | Germination kinetic of Karim (Triticum durum) and Salambô (Triticum aestivum) wheat’s species at low (5°C) and high (45°C) temperatures than transferred ( |
| Table 2: | Water content, peroxidase activity and total protein amount of Karim and Salambô dry seeds |
 | |
Uptake of water by both Karim and Salambô variety is very slow at 5 and 15°C and then it is relatively important at 25, 35 and 45°C.
Results showed in Fig. 3 reveale, very well, that water uptake by wheat seeds (both karim and Salambô species) was done in two phases. The first phase endures 7 h, during it uptake of water is fast at all surrounding temperatures (5, 15, 25, 35 and 45°C). Second phase lasts five hours and was characterized by a slow water uptake. These results were in agreement with those obtained by Bewley and Black (1994) on hard wheat. As shown in Literature data, water uptake by wheat seed comprises two phases: The first independent of surrounding osmotic pressure and corresponds an entry of water into apoplasme.
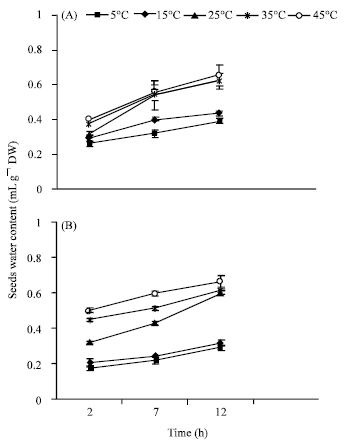 | |
| Fig. 3: | Water uptake by two wheat species (Karim and Salambô) during germination of seeds at different temperatures (5-45°C). Values represent the mean of three repetitions. Confidence intervals are in 5% threshold |
The second phase, very slow than the first and corresponding to a water transit through the cellular membrane of seed. This phase was determined as the osmotic pressure difference between seed and the soil matrix (Allen et al., 2000). At the same, Bellani et al. (2002) have revealed that water uptake during germination of Brassica oleracea contains two phases. The first phase endures 3 h and was characterized by an abrupt increase in fresh weight. The second lasts 24 h, but characterized by a slight increase in weight and came to an end by radicle protrusion (Bellani et al., 2002).
Effect of temperature on reserve mobilization: Effect of heat constraint on seed reserve mobilization during germination was estimated by the residuary dry matter amount of seed. Figure 4 showed that karim cultivar has an average dry matter at the order 48.38 (±0.35) mg. However, dry matter of Salambô cultivar is very low. It is in order of 40.84 (± 0.29) mg. In seeds of both cultivars, residuary dry matter decreases hardly, particularly, between day 2 and day 4 when seeds were germinated at 25°C. Reserve mobilization was very affected by low (5°C) and high (45°C) temperature. This mobilization decreases enormously between 4 and day 6.
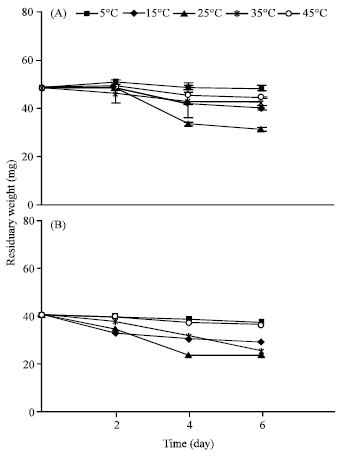 | |
| Fig. 4: | Heat stress influence seed reserves mobilization of two wheat species (Karim and Salambô), assessed by amount of residuary dry matter after 2, 4 and 6 days of germination at the obscurity. Values represent the mean of three repetitions |
These agree with transition from heterotrophic to an autotrophic survival seedlings. Present results confirmed that of (Petruzzelli and Taranto, 1989). These authors showed a decrease in the rate of reserve mobilization and metabolites of seed hard wheat, variety Appulo, to the embryo after 4 days of heat stress application. This drop was associated, also, with loss seed viability (Petruzzelli and Taranto, 1989).
Effect of heat stress on proteins amount: As shown in literature, plants and organisms respond to high temperature stress with the induction of heat shock proteins, HSPs, (Lindquist and Craig, 1988; Vierling, 1991). HSPS are postulated to protect organisms from the damaging effects of heat and others forms of stress.
Obtained results in Fig. 5 show that, during imbibition (first two hours) under high temperature (45°C), protein synthesis was found to be important in both Karim and Salambô species. It may be that sufficient cellular protection had occurred during imbibition (2 h) at 45°C.
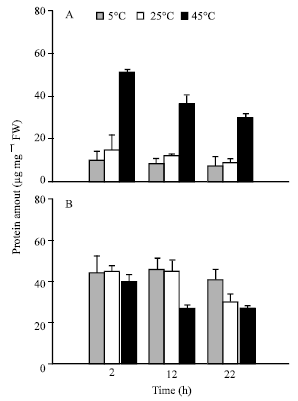 | |
| Fig. 5: | Effect of heat stress on total proteins content of tow wheat seed species during germination (2, 12 and 22 h). Values represent the mean of three repetitions. Confidence intervals are in 5% threshold |
The signature response to heat stress is a decrease in the synthesis of normal proteins, accompanied by an accelerated transcription and translation of a new set of proteins known as heat shock proteins (Perras and Sarhan, 1989). This response is observed when plants are exposed to temperatures at least 5°C above their optimal growing conditions. At 5°C, there is not a markedly change in proteins content during germination. At this stage, a slight decrease in protein amount was observed in control condition (25°C). Present observations in wheat and other reported studies strongly indicate a positive role of HSPs in the protection of the normal metabolic activity during high and low temperature stress.
Effect of temperature on peroxidase activity during seed germination: Preoxidases were involved in growth and development processes (Requelme and Cardemil, 1993). Their activity often varies with external conditions (Castillo, 1992; Yun et al., 1997). In fact, they were considered as biochemical markers of stress. Considering the importance of peroxidases in protection mechanism of plants against constrained conditions (Andreeva, 1991), we were interested to study the effect of heat stress on peroxidase activity of germinated seeds. Peroxidase activity of dry seeds was 31.925 (± 10) and 118.74 (± 6.80) for Karim and Salambô cultivars, respectively (Table 2).
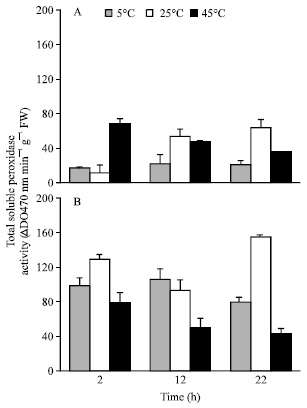 | |
| Fig. 6: | Totals solubles peroxidases activity of two wheat species (Karim and Salambô) during germination at different temperature regimes (5, 25 and 45°C). Values represent the mean of three repetitions. Confidence intervals are in 5% threshold |
This activity varies, during germination, with surrounding temperature. Obtained results in Fig. 6 showed that during imbibition phase, only high temperature (45°C) induce, for Karim cultivar, a strong stimulation of peroxidase activity in comparison with that observed for dry seeds. This stimulation was concomitant to a strong hydratation (Essemine et al., 2006). Viability loss of certain embryos and enzyme activity stimulation were two complimentary hypothesis in favour that 45°C is a constraint temperature for germination.
The result from present study (Essemine et al., 2005 and 2006) show for the first time the protective effect of HSPs on an enzyme during the 2 first hours of imbibition at high temperature (45°C). At 5°C, peroxidase activity of seeds decreased in comparison with that of dry seeds (Fig. 6). This was in agreement with slowing down metabolic process. Present results show, as in salt stress case, that peroxidase activity depends with conditions of germination place (Jbir et al., 2002). Resistance to high temperature and stimulation of peroxidase enzyme in wheat is probably mediated by synthesis of HSPs during mode of life at heat stress, though this might not be the only reason and other mechanisms might exist as well. In fact, plants submitted to environment aggressions developed defensive reactions involving an activation of many genes (Collinge and Salusarenko, 1987; Sturm and Chrispeels, 1990) as a response to various stimulati.
CONCLUSIONS
Germination constitutes a precocious test for researching threshold plants tolerance to heat constraint. Studying wheat seeds behaviour, Karim and Salambô cultivars, during germination at different temperatures (Fig. 1) in comparison with control temperature (25°C) reveales that low temperature reduced germination capacity and delayed germination. However, high temperature affected germination capacity and increased lightly the delay time. The prolonged delay time want to prevent radicle protrusion and defended to obtain seedlings enable to grow properly. These species have an aptitude to germinate at 45°C. The nature of adaptation process and the intensity of the response vary among plants species and among the different cultivars of wheat. The HSPs were found to protect normal proteins from the direct effects of denaturation during heat stress treatment. Jinn et al. (1989) have reported a similar correlation between the thermostabilization of normal proteins and the amount of HSP synthesized. In present studies, peroxidase activity has a crucial role in adaptation to stress.
Understanding the function of HSPs in seeds requires more additional work. More complete characterization of HSP molecular weight by SDS-polyaclrylamide gel electroforesis and biochemical properties are required.
REFERENCES
- Bellani, L.M., M. Guarnieri and A. Scialabba, 2002. Differences in the activity and distribution of peroxidases from three diferent portions of germinate Brassica oleracea seeds. Physiol. Plant, 114: 102-108.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Collinge, D.B. and A.J. Slusarenko, 1987. Plant gene expression in response to pathogens. Plant Mol. Biol., 9: 389-410.
CrossRef - Covell, S., R.H. Ellis, E.H. Roberts and R.J. Summerfield, 1986. The influence of temperature on seed germination rate in grain legumes. I. A comparison of chickpea, lentil, soyabean and cowpea at constant temperatures. J. Exp. Bot., 37: 705-715.
CrossRefDirect Link - Essemine, J., N. Jbir, S. Ammar and S. Bouzid, 2005. Reponses Physiologique Et Biochimique Des Grains De Ble Tendre, Triticum Aestivum, � Une Contrainte Thermique Au Cours De La Germination. Premier Colloque Euromediterraneen en Biologie Vegetale et environnement. Universit� Badji Mokhtar, Annaba, Alg�rie.
- Hampson, C.R. and G.M. Simpson, 1990. Effects of temperature, salt and osmotic potential on early growth of wheat (Triticum aestivum). I. Germination. Can. J. Bot., 68: 524-528.
CrossRefDirect Link - Hardegree, S.P., A.J. Thomas and S.V.V. Steven, 2002. Variability in thermal response to primed and non-primed seeds of Squirreltail (Elymus elymoides (raf.) Swezey and Elymus multisetus (J.G. Smith) M.E. Jones). Ann. Bot., 89: 311-319.
Direct Link - Jbir, N., A. Ayadi, S. Ammar, W. Chaibi and J. Brulfert, 2002. Seed germination of two wheat species differing in their sensitivity to NaCl, in response to salt stress. J. Trace Microprobe Technol., 20: 625-637.
Direct Link - Lindquist, S. and E.A. Craig, 1988. The heat-shock proteins. Ann. Rev. Genet., 22: 631-677.
CrossRefPubMedDirect Link - Murphy, J.B. and T.L. Noland, 1982. Temperature effects on seed imbibition and leakage mediated by viscosity and membranes. Plant Physiol., 69: 428-431.
Direct Link - Perras, M. and F. Sarhan, 1989. Synthesis of freezing tolerance proteins in leaves, crown and roots during cold acclimation of wheat. Plant Physiol., 89: 577-585.
Direct Link - Petruzelli, L. and G. Taranto, 1989. Wheat ageing: The contribution of embryonic and non-embryonic lesions to loss of seed viability. Physiol. Plant., 76: 289-294.
CrossRef - Requelme, A. and L. Cardemil, 1993. Peroxidases in the cell walls of seeds and seedlings of Araucana araucaria. Phytochemistry, 32: 15-20.
Direct Link - Vierling, E., 1991. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 579-620.
CrossRefDirect Link